e.
k+`^`w,q,,'esi^
•?
__
RESEARCH
.
REPORTING SERIES
Research reports
,
of the Office of Research and Development, U.S. Environmental
Protection Agency, have been grouped into
,
nine
`
series. These nine broad cate
gories were established to facilitate further
-
development and application of en
vironmental:technology.. Elimination of, traditional grouping was consciously
planned to foster technology transfer
and
.
maximum interface in related fields.
E
nvironmental Health:Effects Research
Environmental PrOtectiOnTechnologY
Ecological,Research
Environmental
.
?;V:
ocioeconomic
:
?
:•::
?•
Environmental
?
.?
,?
;
?
Studies
.
cientific
r and Technical
.
Assessment Reports (STAR)
nteragency Energy-Environment Research and
. Development
pecial Reports
Mi
scellaneous Reports
his report
_
Has
,
been assigned tp:the
.
ECOLOGICAL RESEARCH
series.ThiS
Series.
escribes
r
esearch on the effects of
pollution on humans,` pplant
and animal spe-:
diO'§,'
. ,:and ,it:atei-ia
.
l
:"
P
r
o61er:rf
.
are :aSsessO
:
fo(thbir
lOng z'::and short-term influ
ences Investigations include formation; transport and
pathway stud es
to deter-,
hth0B*16tS"Pf:PplibtahtrahOtheif...0te:cts:;70is
work provides the
.
techniOal basis',
in the
uatijerrestrialdnd:atriibonoic
nvir
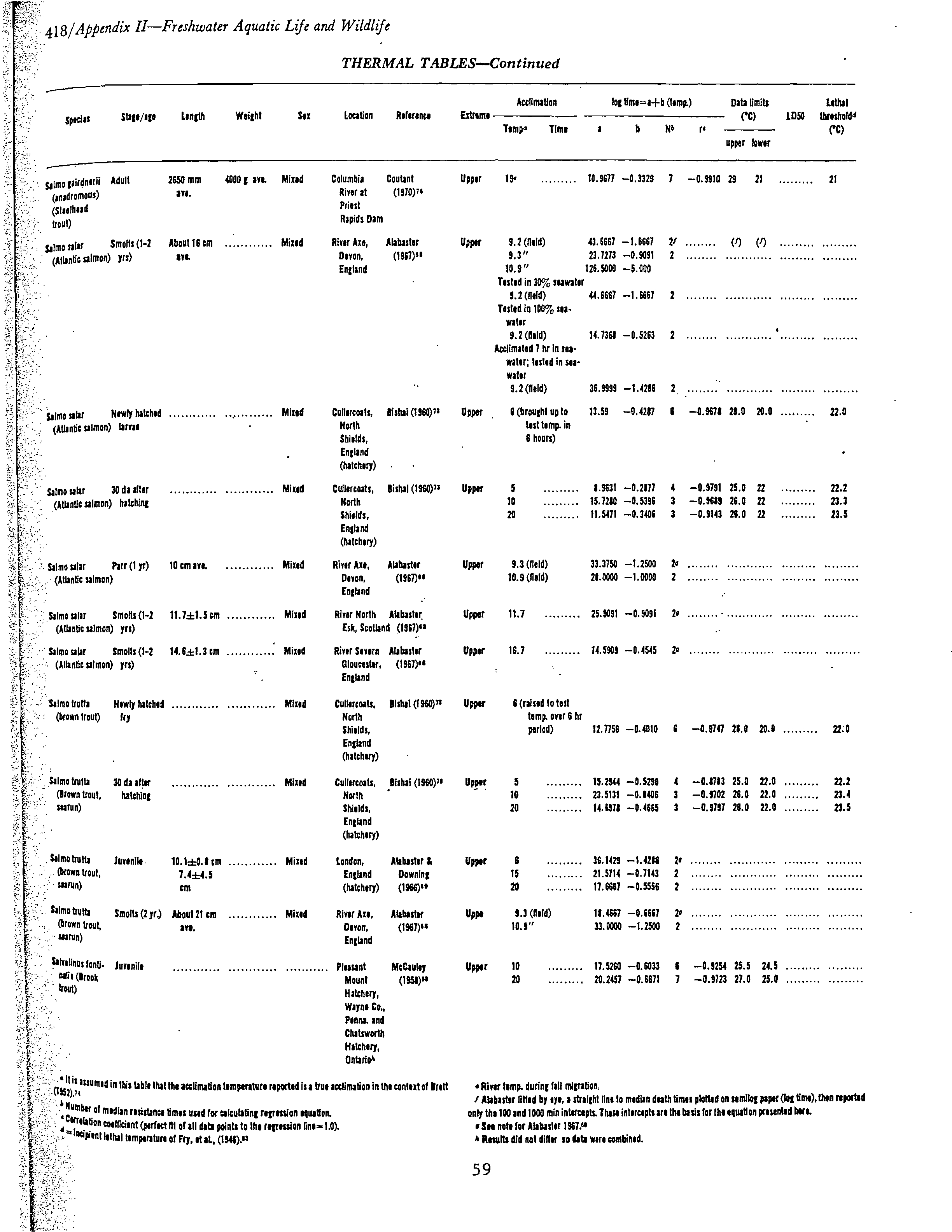
EPA-600/3-77-061
May 1977
TEMPERATURE CRITERIA FOR FRESHWATER FISH:
PROTOCOL AND PROCEDURES
by
William A. Brungs
Bernard R. Jones
Environmental Research Laboratory-Duluth
Duluth, Minnesota 55804
t4,)?
%-4.",,,
CO
?
'''',:
I!
qs
4/to
4',./
..-/c.
1//to?
,,
s„,
6'41
-4*/
<960
?
16/1
d'''
),
ENVIRONMENTAL RESEARCH LABORATORY-DULUTH
?
OA'
„,?Ac.26
U.S.
OFFICE
ENVIRONMENTAL
OF RESEARCH
PROTECTION
AND DEVELOPMENTAGENCY
?
?
..'44`v:4,?e
76
,
09,
t.,
DULUTH, MINNESOTA 55804
DISCLAIMER
This report has been reviewed by the Environmental Research Laboratory—
Duluth, U.S. Environmental Protection Agency, and approved for publication.
Mention of trade names or commercial products does not constitute endorsement
or recommendation for use.
ii
FOREWORD
Our nation's fresh waters are vital for all animals and plants, yet our
diverse uses of water -- for recreation, food, energy, transportation, and
industry -- physically and chemically alter lakes, rivers, and streams. Such
alterations threaten terrestrial organisms, as well as those living in water.
The Environmental Research Laboratory in Duluth, Minnesota, develops methods,
• conducts laboratory and field studies, and extrapolates research findings
--to determine how physical and chemical pollution affects.
aquatic life;
--to assess the effects of ecosystems on pollutants;
--to predict effects of pollutants on large lakes through '
use of models; and
--to measure bioaccumulation of pollutants in aquatic
organisms that are consumed by other animals, including
man.
This report discusses the history, procedures, and derivation of
temperature criteria to protect freshwater fishes and presents numerical
criteria for 34 species. It follows the general philosophical approach
of the National Academy of Sciences and National Academy of Engineering in
their Water Quality Criteria 1972 and is intended to make that philosophy
practically useful.
Donald I. Mount, Ph.D.
Director
Environmental Research. Laboratory
Duluth, Minnesota
iii
ABSTRACT
Temperature criteria for freshwater fish are expressed as mean and
maximum temperatures; means control functions such as embryogenesis, growth,
maturation, and reproductivity, and maxima provide protection for all life
stages-against lethal conditions. These criteria for 34 fish species are
based on numerous field and laboratory studies, and yet for some important
species the data are still insufficient to develop all the necessary
criteria. Fishery managers, power–plant designers, and regulatory agencies
will find these criteria useful in their efforts to protect fishery resource:
iv
CONTENTS
Page
Foreword
?
iii
Abstract
?
iv
Acknowledgments
?
vi
1.
Summary and Conclusions
?
1
2.
Introduction
?
2
3.
The Protocol for Temperature Criteria
?
10
4.
The Procedures for Calculating Numerical Temperature
Criteria for Freshwater Fish
?
13
5.
Examples ? ?
? 20
References
?
25
Appendices
A.
The heat and temperature section from National Academy of..
Sciences and National Academy of Engineering (19731
? 28
B.
The thermal tables from National Academy of Sciences and
National Academy of Engineering (1973)
?
51
C.
Fish temperature data sheets ?
62
ACKNOWLEDGMENTS
We would like to express our appreciation for review of this report to
Dr. Charles C. Coutant (Oak Ridge National Laboratory), Mr. Carlos M.
Fetterolf, Jr. (Great Lakes Fishery Commission), Mr. William L. Klein (Ohio
River Valley Sanitation Commission), and Dr, Donald I. Mount, Dr. Kenneth E. F.
Hokanson and Mr. J. Howard McCormick (Environmental Research Laboratory-
Duluth).
v
i
SECTION 1
SUMMARY AND CONCLUSIONS
The evolution of freshwater temperature criteria has advanced from the
search for a single "magic number" to the generally accepted protocol for
determining mean and maximum numerical criteria based on the protection of
appropriate desirable or important fish species, or both. The philosophy and
protocol of the National Academy of Sciences and National Academy of
,
Engineering (1973) were used to determine criteria for survival, spawning,
embryo development, growth, and gamete maturation for species of freshwater
fish, both warmwater and coldwater species.
The influence that management objectives and selection of species have
on the application of temperature criteria
,
is extremely important, especially
if an inappropriate, but very temperature-sensitive, species is included. In
such a case, unnecessarily restrictive criteria will be derived. Conversely,
if the most sensitive important species is not considered, the resultant
criteria will not be protective.
SECTION 2
INTRODUCTION
This report is intended to be a guide for derivation of temperature
criteria for freshwater fish based on the philosophy and protocol presented
by the National Academy of Sciences and National Academy of Engineering (1973).
It is not an attempt to gather and summarize the literature on thermal effects.
Methods for determination of temperature criteria have evolved and
developed rapidly during the. past 20 years, making possible a vast increase
in basic data on the relationship of temperature to various life stages.
One of the earliest published temperature criteria for freshwater life
was prepared by the Aquatic Life Advisory Committee of the Ohio River Valley
Water Sanitation Commission (ORSANCO) in 1956. These criteria were based on
conditions necessary to maintain a well-rounded fish population and to sustain
production of a harvestable crop in the Ohio River watershed. The committee
recommended that the temperature of the receiving water:
1)
,
Should not be raised above 34° C (93°F) at any place
or at any time;
should not be raised above 23° C (73° F) at any place
or at any time during the months of December through
April; and
3) should not be raised in streams suitable for trout
propagation.
McKee and Wolf (1963) in their discussion of temperature criteria for the
propagation of fish and other aquatic and marine life refer only to the
progress report of ORSANCO's_Aquatic Life Advisory Committee (.1956).
In 1967 the Aquatic Life Advisory Committee of ORSANCO evaluated and
further modified their recommendations for temperature in the Ohio River
watershed. At this time the committee expanded their recommendation of a
93° F (33.9° C) instantaneous temperature at any time or any place to include
a daily mean of 90° F (32.2° C). This, we believe, was one of the first
efforts to recognize the importance of both mean and maximum temperatures
to describe temperature requirements of fishes. The 1967 recommedations also
included:
Maximum temperature during December, January, and February
should be 55° F (12.8° C);
2
2) during the transition months of March, April, October and
November the temperature can be changed gradually by not
more than 7° F (3.9° C);
to maintain trout habitats, stream temperatures should not
exceed 55° F (12.8° C) during the months of October through
May, or exceed 68° F (20.0° C) during the months of June
through September; and
4) insofar as possible the temperature should not be raised
in streams used for natural propagation of trout.
The National Technical Advisory Committee of the Federal Water Pollution
Control Administration presented a report on water quality criteria in 1968
that was to become known as the "Green Book." This large committee included
many of the members of ORSANCO's Aquatic Life Advisory Committee. The committee
members recognized that aquatic organisms might be able to endure a high
temperature for a few hours that could not be endured for a period of days.
They also acknowledged that no single temperature requirement could be applied
to the United States as a whole, or even to one state, and that the requirements
must be closely related to each body of water and its fish populations. Other
important conditions for temperature requirements were that (1) a seasonal cycle
must be retained, (2) the changes in temperature must be gradual, and (3) the
temperature reached must not be so high or so low as to damage or alter the
composition of the desired population. These conditions led to an approach to
criteria development different from earlier ones. A temperature increment
based on the natural water temperature was believed to be more appropriate
than an unvarying number. The use of an increment requires a knowledge of
the natural temperature conditions of the water in question, and the size of
the increment that can be tolerated by the desirable species.
The National Technical Advisory Committee (1968, p. 42) recommended:
"To maintain a well–rounded population of warmwater fishes .... heat
- should not be added to a stream in excess of the amount that will
raise the temperature of the water (at the expected minimum daily
flow for that month) more than 5° F."
A casual reading of this requirement resulted in the unintended generalization
that the acceptable temperature rise in warmwater fish streams was 5° F (2.8°
C). This generalization was incorrect! Upon more careful reading the key
word "amount" of heat and the key phrase "minimum daily flow for that month"
clarify the erroneousness of the generalization. In fact, a 5° F (2.8° C)
rise in temperature could only be acceptable under low flow conditions for a
particular month and any increase in flow would result in a reduced increment
of temperature rise since the amount of heat added could not be increased.
For lakes and reservoirs the temperature rise limitation was 3° F (1.7° C)
based "on the monthly average of the maximum daily temperature."
In trout and salmon waters the recommendations were that "inland trout
streams, headwaters of salmon streams, trout and salmon lakes, and reservoirs
containing salmonids should not be warmed," that "no heated effluents should
3
be discharged in the vicinity of spawning areas," and that "in lakes and
reservoirs, the temperature of the hypolimnion should not be raised more
than 3° F (1.7° C)." For other locations the recommended incremental rise
was 5° F (2.8° C) again based on the minimum expected flow for that month.
An important additional recommendation is summarized in the following
table in which provisional maximum temperatures were recommended for various
fish species and their associated biota (from FWPCA National Technical Advisory
Committee, 1968).
PROVISIONAL MAXIMUM TEMPERATURES RECOMMENDED AS
COMPATIBLE WITH THE WELL-BEING OF VARIOUS SPECIES
OF FISH AND THEIR ASSOCIATED BIOTA
93 F: Growth of catfish, gar, white or yellow bass, spotted
bass, buffalo, carpsucker, threadfin shad, and
gizzard
shad.
90 F: Growth of largemouth bass, drum, bluegill, and crappie.
84 F: Growth of pike, perch, walleye, smallmouth bass, and
sauger.
80 F; Spawning and egg development of catfish, buffalo, thread-
fin shad, and gizzard shad.
75 F: Spawning and egg development of largemouth bass, white,
yellow, and spotted bass.
68 F; Growth or migration routes of salmonids and for egg
development of perch and smallmouth bass.
55 F: Spawning and egg development of salmon and trout (other
than lake trout).
48 F: Spawning and egg development of lake trout, walleye,
northern pike, sauger, and Atlantic salmon.
NOTE: Recommended temperatures for other species, not listed
above, may be established if and when necessary
information becomes available.
These recommendations represent one of the significant early efforts to base
telaperature criteria on the realistic approach of species and community
requirements and take into account the significant biological factors of
spawning, embryo development, growth, and survival.
?
•
4
The Federal Water Pollution Control Administration (1969a) recommended
revisions in water quality criteria for aquatic life relative to the Main
Stem of the Ohio River. These recommendations were presented to ORSANCO's
Engineering Committee and were based on the temperature requirements of
important Ohio River fishes including largemouth bass, smallmouth bass, white
bass, sauger, channel catfish, emerald shiner, freshwater drum, golden
redhorse, white sucker, and buffalo (species was not indicated). Temperature
requirements for survival, activity, final preferred temperature, reproduction,
and growth were considered. The recommended criteria were:
1.
"The water temperatures shall not exceed 90° F
(32.2° C) at any time or any place, and a
maximum hourly average value of 86° F (30° C)
shall not be exceeded."
2.
"The temperature shall not exceed the
temperature values expressed on the following
table:"
AQUATIC LIFE TABLEa
Daily mean
(°
F).
Hourly maximum
(
Q
F)
December-February
48
55
Early March
50
56
Late March
52
58
Early April
55
60
Late April
58
62
Early May
62
64
Late May
68
72
Early June
75
79
Late June
78
82
July-September
82
86
October
75
82
November
65
72
a
From: Federal Water Pollution Control Administration
(1969a).
5
The principal limiting fish species considered in developing these criteria
was the sauger, the most temperature sensitive of the important Ohio River
fishes. A second set of criteria (Federal Water Pollution Control
Administration, 1969b) considered less temperature-sensitive species, and the
criteria for mean temperatures were higher. The daily mean in July and
September was 84° F (28.9° C). In addition, a third set of criteria was
developed that was not designed to protect the smallmouth bass, emerald
shiner, golden redhorse, or the white sucker. The July-to-September daily
mean temperature criterion was 86° F (30° C).
The significance of the 1969 Ohio River criteria was that they were
species dependent and that subsequently the criteria would probably be based
upon a single species or a related group of species. Therefore, it is
extremely important to select properly the species that are important otherwise
the criteria will be unnecessarily restrictive. For example, if yellow perch
is an extremely rare species in a water body and is the most temperature-
sensitive species, it probably would be unreasonable to establish temperature
criteria for this species as part of the regulatory mechanism.
In 1970 ORSANCO established new temperature standards that incorporated
the recommendations for temperature criteria of the Federal Water Pollution
Control Administration (1969a, 1969b)
.
and the concept of limiting the amount
of heat that would be added (National Technical Advisory Committee, 1968).
The following is the complete text of that standard:
" All cooling water from municipalities or political
subdivisions, public or private institutions, or
installations, or corporations discharged or
permitted to flow into
,
the Ohio River from the point
of confluence of the Allegheny and Monongahela Rivers
at Pittsburgh, Pennsylvania, designated as
Ohio River
mile point 0.0 to Cairo Point, Illinois, located at
the confluence of the Ohio and Mississippi Rivers, and
being 981.0 miles downstream from Pittsburgh, Pennsylvania,
shall be so regulated or controlled as to provide for
reduction of heat content to such degree that the aggregate
heat-discharge rate from the municipality, subdivision,
institution, installation or corporation, as calculated on
the basis of discharge volume and temperature. differential
(temperature of discharge minus upstream river temperature)
does not exceed the amount calculated by the following
formula, provided, however, that in no case shall the
aggregate heat-discharge rate be of such magnitude as will
result in a calculated increase in river temperature of
more than 5 degrees F:
Allowable heat-discharge rate (Btu/sec) = 62.4 X
river flow (CFS) X (T
a
- T
r
) X 90%
6
Where:
T
a
= Allowable maximum temperature (deg. F.)
in the river as specified in the following
table:
January
50
July
89
February
50
August
89.
March
60
September
87
April
70
October
78
May
80.
November
70
June
87
December
57
T
r
= River temperature (daily average in deg. F.I
upstream from the discharge
River flow = measured flow but not less than
critical flow values specified in
the following table:
River reach
?
Critical
flow
From
?
in cfsa
Pittsburgh, Penn.
(mi.
O.0) ?
Willow Is. Dam (161.7)
?
6,500.
Willow Is. Dam (161.7)
?
Gallipolis Dam (279.21
?
7,400
Gallipolis Dam (279.21?
Meldahl Dam (436.21
?
9,700
Meldahl Dam (436.2)?
McAlpine Dam (605.8)
?
11,900
McAlpine Dam (605.8)
?
Uniontown Dam (846.0)
?
14,200
Uniontown Dam (846.0)
?
Smithland Dam (918.51
?
19,500
Smithland Dam (918.5)
?
Cairo Point (981.0)
?
48,100
a
Minimum daily flow once in ten years.
7
Although the numerical criteria for January through December are higher
than those recommended by the Federal Water Pollution Control Administration,
they are only used to calculate the amount of heat that can be added at the
"minimum daily flow once in ten years." Additional flow would result in
lower maxima since no additional heat could be added. There was also the
increase of 5° F (2.8° C) limit that could be more stringent than the maximum
temperature limit.
The next important step in the evolution of thought on temperature
criteria was Water Quality Criteria 1972 (NAS/NAE, 1973), which is becoming
known as the "Blue Book," because of its comparability to the Green Book (FWPCA
National Technical Advisory Committee, 1968). The Blue Book is the report of
the Committee on Water Quality Criteria of the National Academy of Sciences at
the request of and funded by the U.S. Environmental Protection Agency (EPA).
The heat and temperature section, with its recommendations and appendix data,
was authored by Dr. Charles Coutant of the Oak Ridge National Laboratory. These
materials are reproduced in full in Appendix A and Appendix B in this report.
A discussion and description of the Blue Book temperature criteria will be
found later in this report.
The Federal Water Pollution Control Act Amendments of 1972 (Public
Law 92-500) contain a section [304 (al
C11]
that requires that the
administrator of the EPA "after
. consultation with appropriate Federal and
State agencies and other interested persons, shall develop and publish,
within one year after enactment of this title (and from time to time
thereafter revise) criteria for water'quality accurately reflecting the
latest scientific knowledge (A), on the kind and extent of
ell
identifiable
effects on health and welfare including, but not limited tb, plankton,
fish, shellfish, wildlife, plant life, shorelines, beaches, esthetics, and
recreation which may be expected from the
.
presence of pollutants in any
body of water, including ground water; (B) on the concentration and dispersal
of pollutants or their byproducts, through biological, physical, and
chemical processes; and (C) on the effects of pollutants on biological
community diversity, productivity, and stability, including information on
the factors affecting rates of eutrophication and rates of organic and
inorganic sedimentation for varying types of receiving waters."
The U.S. Environmental Protection Agency (1976) has published Quality
Criteria for Water as a response to the Section 304(a)(1) requirements of
PL 92-500. That approach to-the determination of temperature criteria for
freshwater fish is essentially the same as the approach recommended in the
Blue Book (NAS/NAE, 1973). The EPA criteria report on temperature included
numerical criteria for freshwater fish species and a nomograph for winter
temperature criteria. These detailed criteria were developed according
to the protocol in the Blue Book, and the procedures used to develop those
criteria will be discussed in detail in this report.
The Great Lakes Water Quality Agreement (1972) between the United States
of America and Canada was signed in 1972 and contained a specific water
quality objective for temperature. It states that "There should be no change
that would adversely affect any local or general use of these waters." The
8
International Joint Commission was designated to assist in the implementation
of this agreement and to give advice and recommendations to both countries
on specific water quality objectives. The International Joint Commission
committees assigned the responsibility of developing these objectives have
recommended temperature objectives for the Great Lakes based on the "Blue
Book" approach and are in the process of refining and completing those
objectives for consideration by the commission before submission to the two
countries for implementation.
A
se
SECTION 3
THE PROTOCOL FOR TEMPERATURE CRITERIA
This section is a synthesis of concepts
.
and definitions from Fry et al.
(1942, 1946), Brett (1952, 1956), and the NAS/NAE (1973).
The lethal threshold temperatures are those temperatures at which 50
percent of a sample of individuals would survive indefinitely after acclimation
at some other temperature. The majority of the published literature (Appendix
B) is calculated on the basis of 50 percent survival. These lethal thresholds
are commonly referred to as incipient lethal temperatures. Since organisms
can be lethally stressed by both rising and falling temperatures, there are
upper incipient lethal temperatures and lower incipient lethal temperatures.
These are determined by removing the organisms from a temperature to which
they are acclimated and instantly placing them in a series of other temperatures
that will typically result in a range in survival from 100 to 0 percent.
Acclimation can require up to 4 weeks, depending upon the magnitude of the
difference between the temperature when the fish were obtained and the desired
acclimation temperature, In general, experiments to determine incipient
lethal temperatures should extend until all the organisms in any test chamber
are dead or sufficient time has elapsed for death to have occurred, The
ultimate upper incipient lethal temperature is that beyond which no increase
in lethal temperature is accomplished by further increase in acclimation
temperature. For most freshwater fish species in temperate latitudes the
lower incipient lethal temperatures will usually end at 0° C, being limited
by the freezing point of water. However, for some important species, such as
threadfish shad in freshwater and menhaden in seawater, the lower incipient
lethal temperature is higher than 0° C,
As indicated earlier, the heat and temperature section of the Blue Book
and its associated appendix data and references have been reproduced in this
report as Appendix A and Appendix B. The following discussion will briefly
summarize the various types of criteria and provide some additional insight
into the development of numerical criteria. The Blue Book (Appendix A)
also describes in detail the use of the criteria in relation to entrainment.
MAXIMUM WEEKLY AVERAGE TEMPERATURE
For practical reasons the maximum weekly average temperature (MWAT) is
the mathematical mean of multiple, equally spaced, daily temperatures over a
7-day consecutive period.
10
For Growth
To maintain growth of aquatic organisms at rates necessary for sustaining
actively growing and reproducing populations, the MWAT in the zone normally
inhabit
ed
by the species at the season should not exceed the optimum temperature
plus one-third of the range between the optimum temperature and the ultimate
upper incipient lethal temperature of the species:
ultimate upper incipient?
optimum
?MAT for growth = optimum temperature + lethal temperature
?temperature
3
The optimum temperature is assumed to be the optimum for growth, but other
physiological optima may be used in the absence of growth data. The MWAT need
not apply to accepted mixing zones and must be applied with adequate under-
standing of the normal seasonal distribution of the important species.
For Reproduction
The MWAT for reproduction must consider several factors such as gonad
growth and gamete maturation, potential blocking of spawning migrations,
spawning itself, timing and synchrony with cyclic food sources, and normal
patterns of gradual temperature changes throughout the year. The protection
of reproductive activity must take into account months during which these
processes normally occur in specific water bodies for which criteria are
being developed.
For Winter Survival
The MWAT for fish survival during winter will apply in any area in which
fish could congregate and would include areas such as unscreened discharge
channels. This temperature limit should not exceed the acclimation, or plume,
temperature (minus a 3.6° F (2,0° C) safety factor) that raises the lower
lethal threshold temperature above the normal ambient water temperature for
that season. This criterion will provide protection from fish kills caused
by rapid changes
- in temperature due to plant shutdown or movement of fish
from a heated plume to ambient temperature.
SHORT-TERM EXPOSURE TO EXTREME TEMPERATURE
It is well established that fish can withstand short exposure to temperatures
higher than those acceptable for reproduction and growth without significiant
adverse effects. These exposures should not be too lengthy or frequent or the
species could be adversely affected. The length of time that 50 percent of a
population will survive temperature above the incipient lethal temperature can
be calculated from the following regression equation:
log time (min) = a + b (temperature in °C);
or
temperature (°C) = (log time (min) - a)/b.
The constants "a" and "b" are for intercept and slope and will be discussed
later. Since this equation is based on 50 percent survival, a 3.6° F (2.0° Cl
reduction in the upper incipient lethal temperature will provide the safety
factor to assure no deaths.
For those interested in more detail or the rationale for these general
criteria, Appendices A and B should be read thoroughly. In addition, Appendix
A contains a fine discussion of a procedure to evaluate the potential thermal
impact of aquatic organisms entrained in cooling water or the discharge
plume, or both.
12
SECTION 4
THE PROCEDURES FOR CALCULATING NUMERICAL
TEMPERATURE CRITERIA FOR FRESHWATER FISH
MAXIMUM WEEKLY AVERAGE TEMPERATURE
The necessary minimum data for the determination of this criterion are
the physiological optimum temperature and the ultimate upper incipient lethal
temperature, The latter temperature represents the "breaking point" between
the highest temperatures to which an animal can be acclimated and the lowest
of the extreme upper temperatures that will kill the warm-acclimated organism.
Physiological optima can be based on performance, metabolic rate, temperature
preference, growth, natural distribution, or tolerance. However, the most
sensitive function seems to be growth rate, which appears to be an integrator
of all physiological responses of an organism, In the absence of data on
optimum growth, the use of an optimum for a more specific function related to
activity and metabolism may be more desirable than not developing any growth
criterion at all,
The MWAT's for growth were calculated for fish species for which appropriate
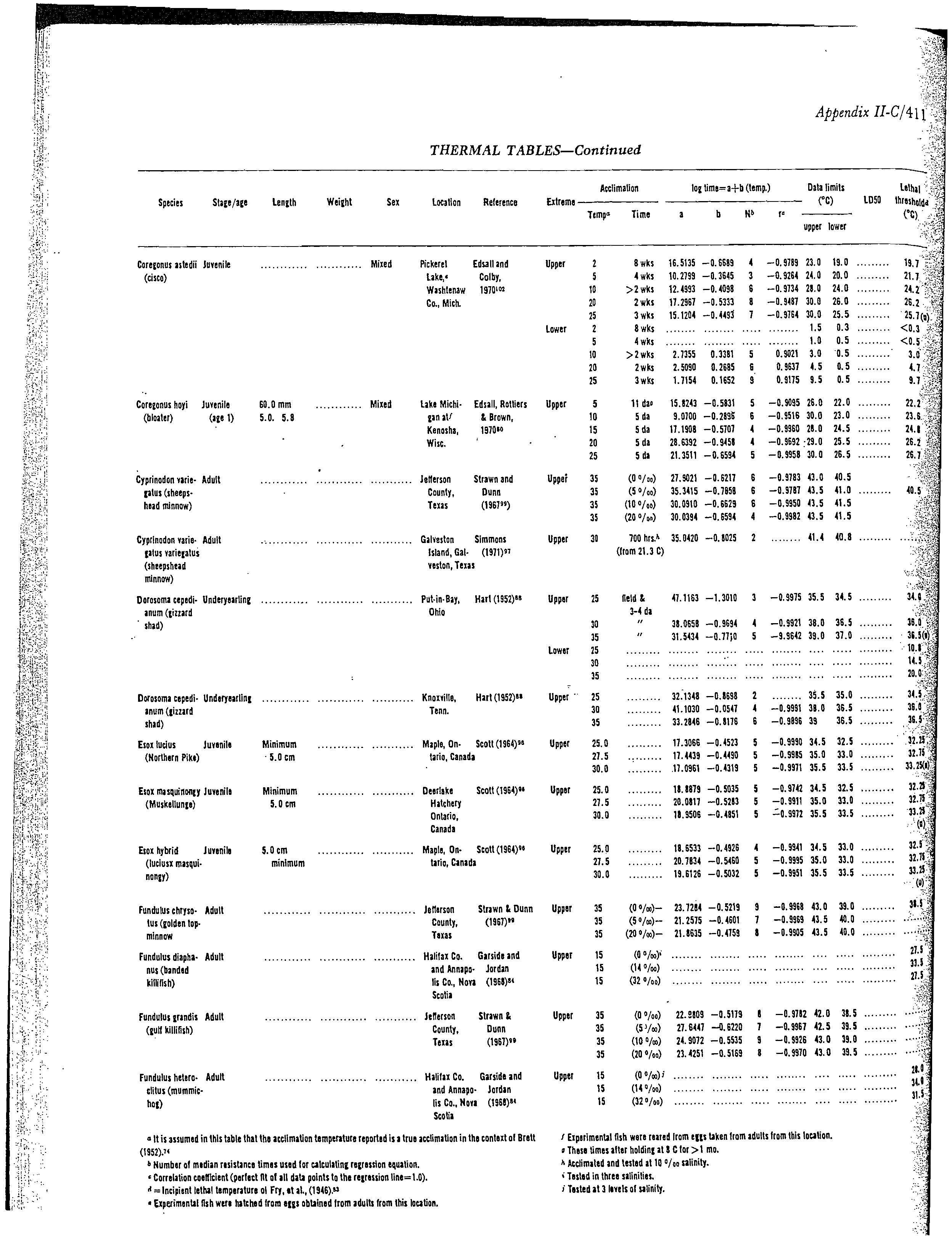
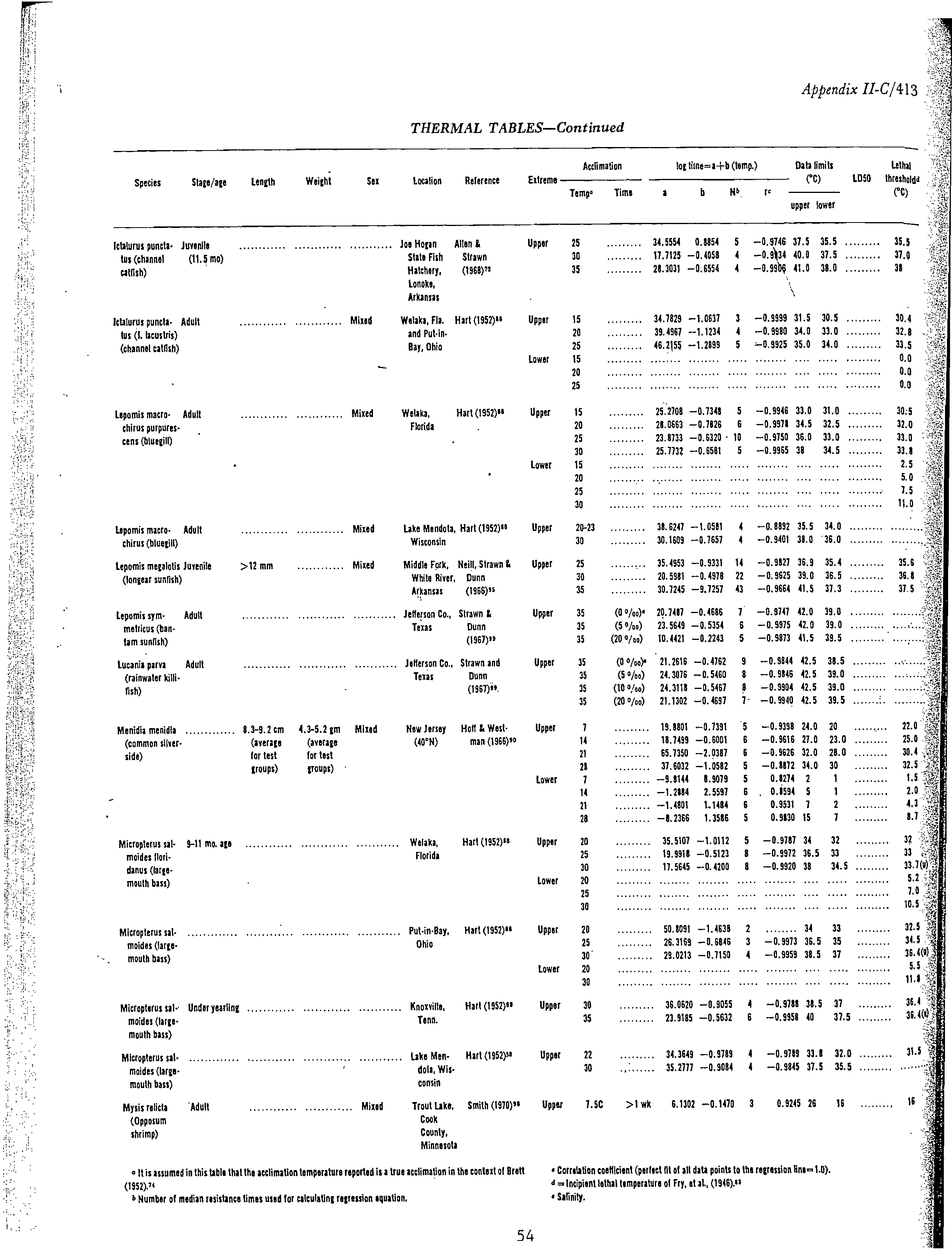
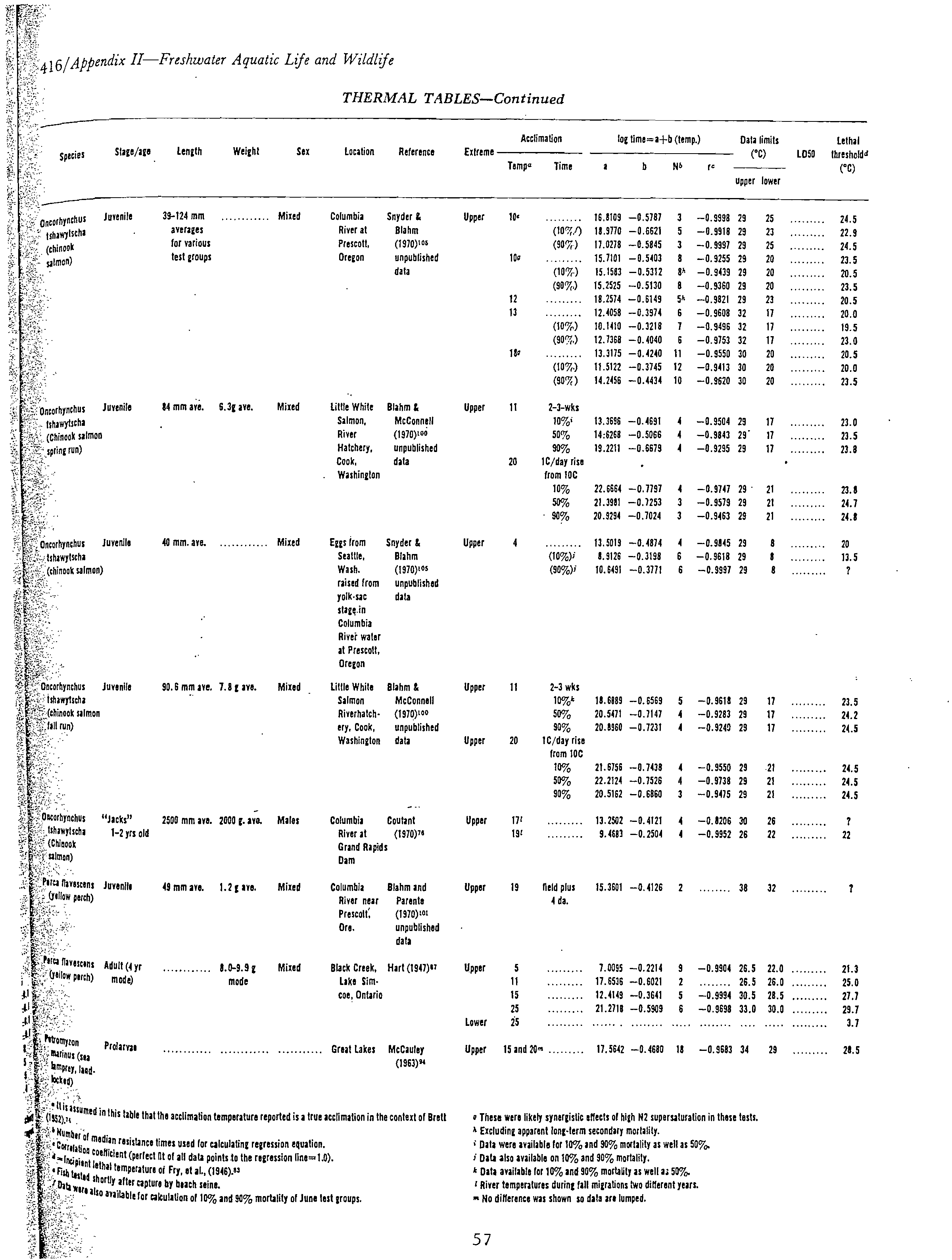
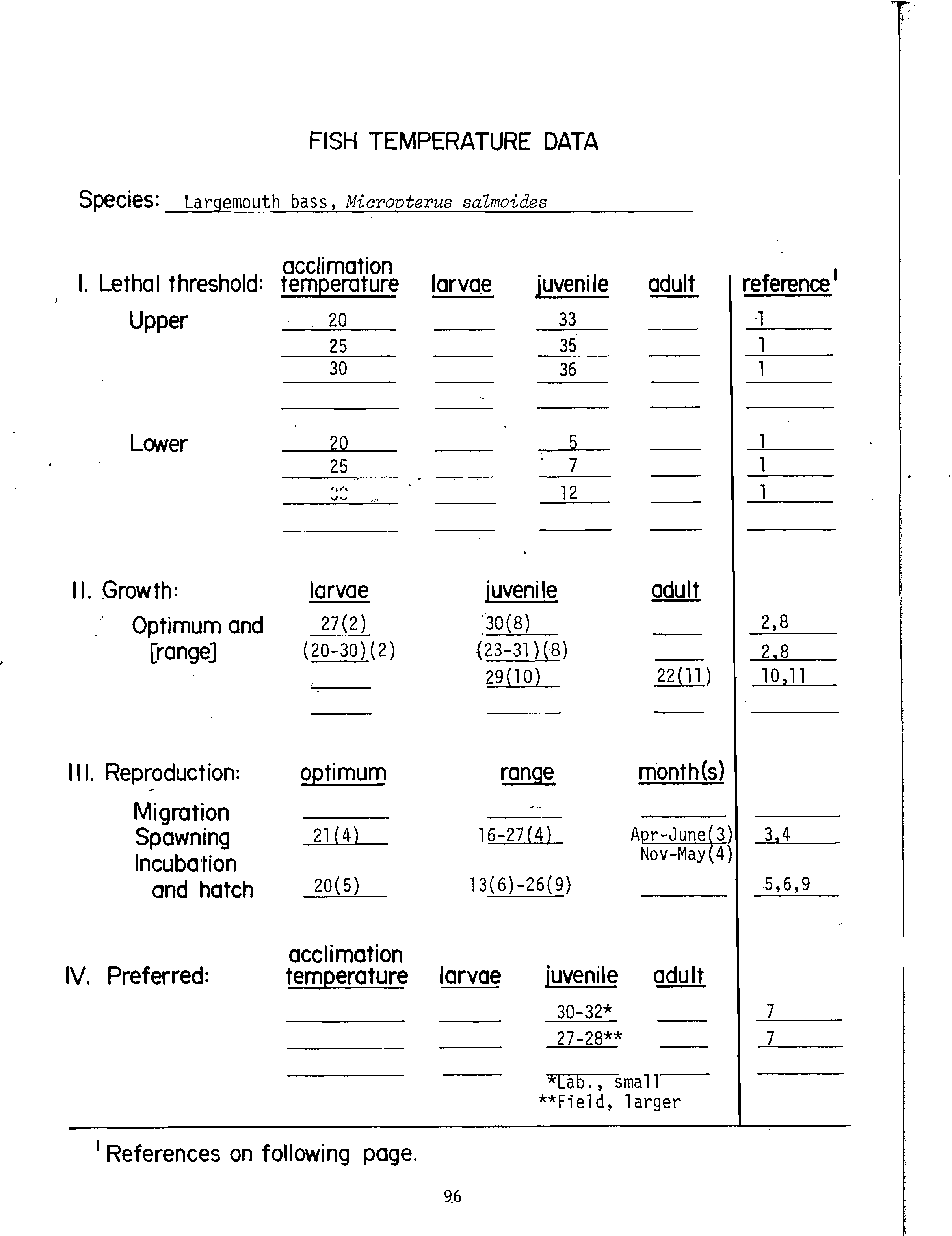
data were available (Table 1). These data were obtained from the fish temperature
data in Appendix C. These data sheets contain the majority of thermal effects
data for about 34 species of freshwater fish and the sources of the data. Some
subjectivity is inevitable and necessary because of variability in published
data resulting from differences in age, day length, feeding regime, or methodology.
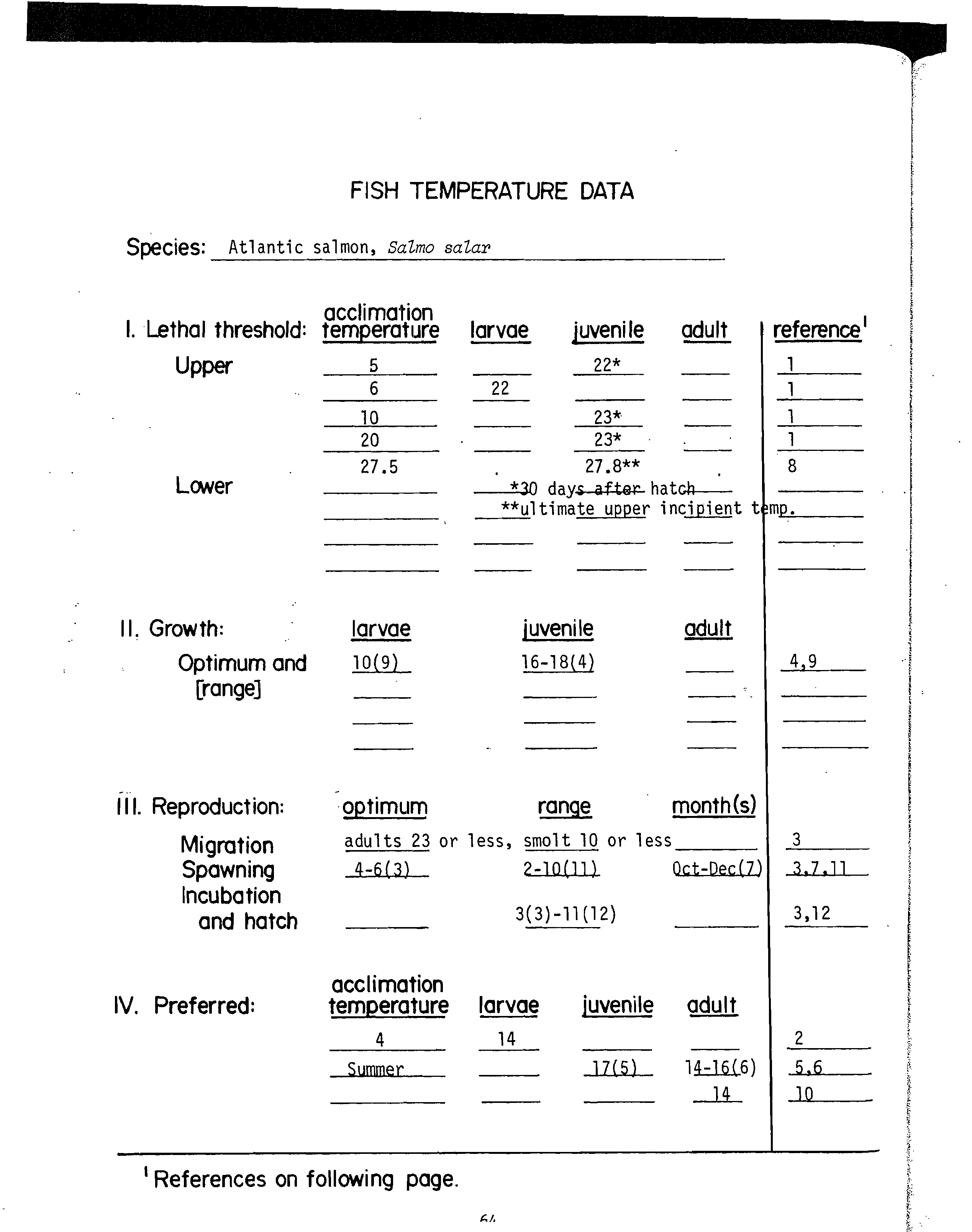
For example, the-data sheet for channel catfish (Appendix C) includes four
temperature ranges for optimum growth based on three published papers.' It would
be. more appropriate to use data for growth of juveniles and adults rather than
larvae. The middle of each range for juvenile channel catfish growth is 29° and
30° C. In this instance 29° C is judged the best estimate of the optimum. The
highest incipient lethal temperature (that would approximate the ultimate
incipient lethal temperature) appearing in Appendix C is 38° C. By using the
previous formula for the MWAT for growth, we obtain
29° C +
(38-29°
3
C)
= 32° C.
The temperature criterion for the MWAT for growth of channel catfish would be
32° C (as appears in Table 1).
13
TABLE 1. TEMPERATURE CRITERIA FOR GROWTH AND SURVIVAL OF SHORT EXPOSURES
(24 HR) OF JUVENILE AND ADULT FISH DURING THE SUMMER (° C (° F))
Species
Maximum weekly average
temperature for growth
s
Maximum temperature for
survival of short exposure
Alewife
--
Atlantic salmon
20 (68)
23 .(73)
Bigmouth buffalo
--
Black crappie
27 (81)
Bluegill
32 (90)
35 (95)
Brook trout
19 (66)
24
(75)
Brown bullhead
' --
Brown trout
17
(63)
24
(75)
Carp
--
Channel catfish
32
(90)
35 (95)
Coho salmon
18
(64)
24
(75)
Emerald shiner
30
(86)
Fathead minnow
Freshwater drum
lake herring (nista)
--
17 (63)
t
25
(77)
Lake whitefish
-7
Lake trout
Largemouth bass
32 (90)
34 (93)
Northern pike
28 (82)
,
?30
(86)
Pumpkinseed
--
Rainbow smelt
Rainbow trout
19
(66)
24 (7S)
Sauger
25
(77)
Smallmouth bass
29 (84)
Smallmouth buffalo
-
Sockeye salmon
16
(64)
22 (72)
Striped bass
Threadfin shad
Walleye
25
(77)
White bass
White crappie
28 (82)
White perch
White sucker
28 (82)t
Yellow perch
29 (84)
a
Calculated according to equation:
maximum weekly average temperature for growth optimum for growth
+ (1/3) (ultimate incipient lethal temperature - optimum for growth).
b
Based on: temperature (. C)
?
(log time (sin) - a)/b - 2
a
C, acclimation
at the maximum weekly average temperature for summer growth, and data in
Appendix B.
c
ilased on data for larvae.
14
SHORT-TER
M
MAXIMUM DURING GROWTH SEASON
In addition to the MWAT, maximum temperature for short exposure will
protect against potential lethal effects. We have to assume that the incipient
lethal temperature data reflecting 50 percent survival necessary for this
calculation would be based on an acclimation temperature near the MWAT for
growth. Therefore, using the data in Appendix B for the channel catfish, we
find four possible data choices near the MWAT of 32° C (again it is preferable
to use data on juveniles or adults):
Acclimation temperature
30? 32.1736
?
-0.7811
34
?
26.4204?
-0.6149
30
? 17,7125
?
-0.4058
35
?
28.3031?
-0.6554'
The formula for calculating the maximum for short exposure is:
temperature (°C) = (log time (min) - a)/b
To solve the equation we must select a maximum time limitation on this
maximum for short exposure, Since the MWAT is a weekly mean temperature
an appropriate length of time for this limitation for short exposure would
be 24 hr without risking violation of the MWAT.
Since the time is fixed at 24 hr (1,440 min), we need to solve for
temperature by using, for example, the above acclimation temperature of 30° C
for which a = 32.1736 and b = -0.7811,
temperature (° C)
log 1,440 -a
3.1584?
-32.1736
?
-29.0152
temperature
-0.7811
?
(°
-0.7811
C)
?
37,146
on solving for each of the four data points we obtain 37,1°, 37.8°, 35.9°, and
38.4° C. The average would be 37.3° C, and after subtracting the 2° C safety
factor to provide 100 percent survival, the short-term maximum for channel
catfish would be 35° C as appears in Table 1.
MAXIMUM WEEKLY AVERAGE TEMPERATURE FOR SPAWNING
From the data sheets in Apendix C one would use either the optimum
t
emperature for -spawning or, if that is_not available, the middle of the range
of
te
mperatures for spawning. Again, if
we
Use the channel catfish 'as an example,
the MWAT for spawning would be
.
27° C (Table 2). Since spawning may occur over
a period of a few weeks or months in a particular water body and only a MWAT
for
o
ptimum spawning is estimated, it would be logical to use that optimum for
the median time of the spawning season. The MWAT for the next earlier month
(° C)
a
b
15
TABLE 2. TEMPERATURE CRITERIA FOR SPAWNING AND EMBRYO SURVIVAL OF
SHORT EXPOSURES DURING THE SPAWNING SEASON (° C
(
0
F))
Species?
'
Maximum weekly average
temperature for spawning°
Maximum temperature for
embryo survivalb
Alewife
22?(72)
28?(82)c
Atlantic salmon
5?(41)
11?
(52)
Bigmouth buffalo
17?(63)
27?(81)c
Black crappie
17?
(63)
• 20?
(68)c
Bluegill
25?
(77)
?
•
34
?
(93)
Brook trout
9?
(48)
13?
(55)
Brown bullhead
24?(75)
27?
(81)
Brown trout
8?
(46)
15?
(59)
Carp
21'(70)
33?
(91)
Channel catfish
27?(81)
29
?
(84)c
Coho salmon
10?
(SO)
13?
(55)c
Emerald shiner
24?
(75)
28?
(82)c
Fathead minnow
24?
(75)
30?
(86)
Freshwater drum
21?
(70)
26?
(79)
Lake herring (disco)
3
?
(37)
8?(46)
Lake whitefish
5?(41)
10?
(50)c
Lake trout
9?
(48)
14?
(57)
Largemouth bass
21?
(70)
27?
(81)c
Northern pike
11?(52)
.
?19?(66)
Fumpkinieed
25?(77)
29?
(84)C
Rainbow smelt
8?(46)
15?
(59)
Rainbow trout
9?
(48)
13?
(55)
Sanger
12?
(54)
18?(64)
Smallmouth bass
17?
(63)
23?
(73)c
Smallrnouth buffalo
21?
(70)
28?
(82)c
Sockeye salmon
10?(50)
13?
(55)
Striped bass
18
?(64)
24?(75)
Throadfin shad
19
?
(66)
34?
(93)
Walleye
8?(46)
17?(63)c
White bass
17
?(63)
26?
(79)
White crappie
18?
(64)
23?
(73)
White perch
15?
(59)
20?
(66)c
White sucker
10?
(50)
20?(68)
Yellow perch
.12?
(54)
20?(68)
4
The optimum or mean of the range of spawning temperatures reported for the
species.
b The upper temperature for successful incubation and hatching reported for
the species.
c Upper temperature for spawning.
16
could approximate the lower temperature of the range in spawning temperature,
and the MWAT for the last month of a 3-month spawning season could approximate
the upper temperature for the range. For example, if the channel catfish
spawned from April to June the MWAT's for the 3 months would be approximately
21°, 27°, and 29° C. For fall spawning fish species the pattern or sequence
of temperatures would be reversed because of naturally declining temperatures
during their spawning season.
SHORT-TERM MAXIMUM DURING SPAWNING SEASON
If spawning season maxima could be determined in the same manner as those
for the growing season, we would be using the time-temperature equation and
the Appendix B data as before. However, growing season data are based usually
on survival of juvenile and adult individuals. Egg-incubation temperature
requirements are more restrictive (lower), and this biological process would
not be protected by maxima based on data for juvenile and adult fish. Also,
spawning itself could be prematurely stopped if those maxima were achieved.
For most species the maximum spawning temperature approximates the maximum
successful incubation temperature. Consequently, the short-term maximum
temperature should preferably be based on maximum incubation temperature for
successful embryo survival, but the maximum temperature for spawning is an
acceptable alternative. In fact, the higher of the two is probably the
preferred choice as variability in available data has shown discrepancies in
this relationship for some species.
For the channel catfish (Appendix C) the maximum reported incubation
temperature is 28° C, and the maximum reported spawning temperature is 29° C.
Therefore, the best estimate of the short-term survival of embryos would be
29° C (Table 2).-
MAXIMUM WEEKLY AVERAGE TEMPERATURE FOR WINTER
•
As discussed earlier the MWAT for winter is designed usually to prevent
fish deaths in the event the water temperature drops rapidly to an ambient
condition. Such a temperature drop could occur as the result of a power-plant
shutdown or a movement
of
the fish itself. These MWAT's are meant to apply
wherever fish can congregate, even if that is within the mixing zone.
Yellow perch require a long chill period during the winter for optimum
egg maturation and spawning (Appendix A). However, protection of this species
would be outside the mixing zone. In addition, the embryos of fall spawning
fish such as trout, salmon, and other related species such as cisco require
low incubation temperatures. For these species also the MAT during winter
would have to consider embryo survival, but again, this would be outside the
mixing zone, The mixing zone, as used in this report, is that area adjacent
to the discharge in which receiving system water quality standards do not
apply; a thermal plume therefore is not a mixing zone.
With these exceptions in mind, it is unlikely that any signficant
effects on fish populations would occur as long as death was prevented.
17
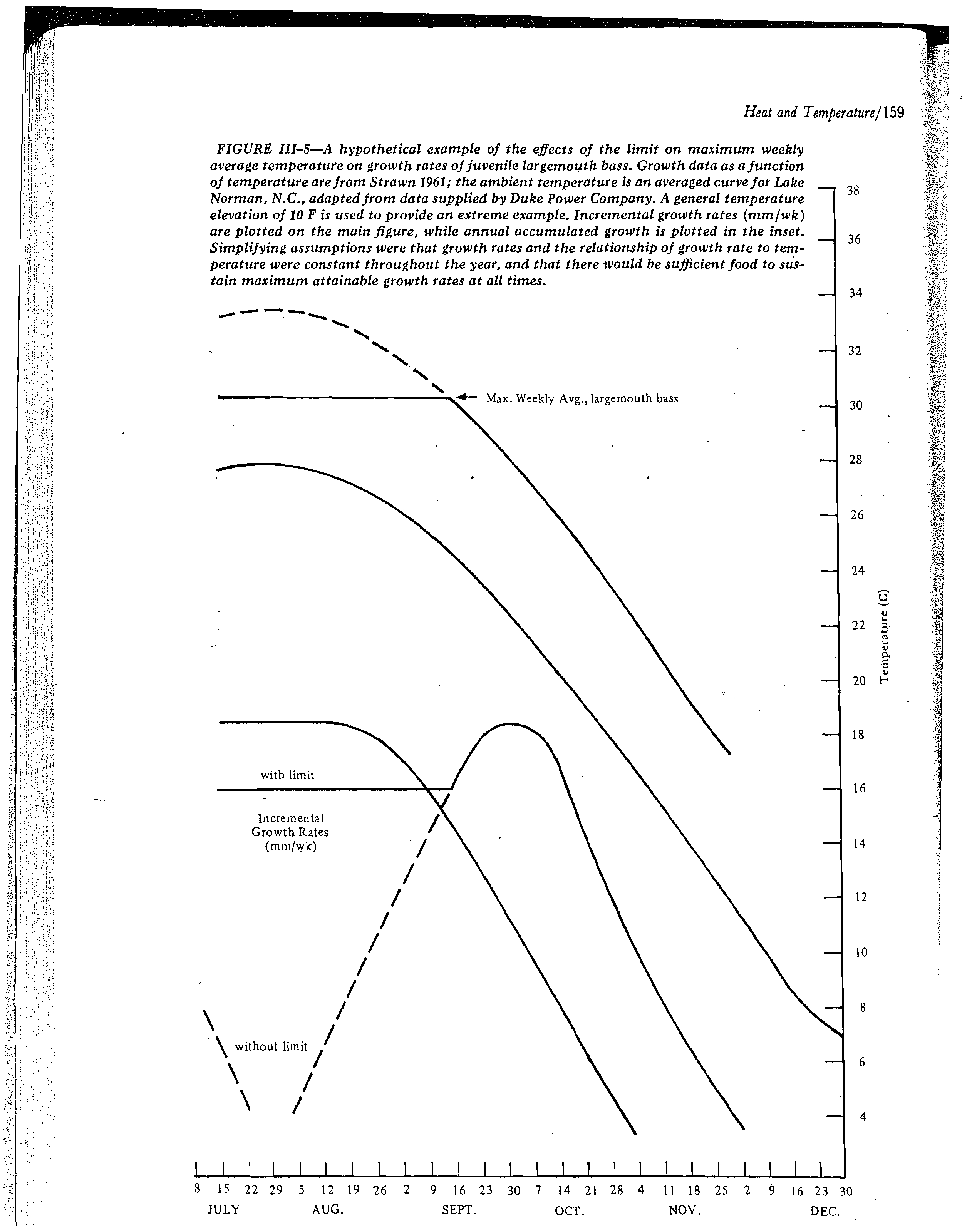
In many instances growth could be enhanced by controlled winter heat addition,
but inadequate food may result in poor condition of the fish.
There are fewer data for lower incipient lethal temperatures than for
the previously discussed upper incipient lethal temperatures. Appendix B
contains lower incipient lethal temperature data for only about 20 freshwater
fish species, less than half of which are listed in Tables 1 and 2. Consequent:
the available data were combined to calculate a regression line (Figure 1)
which gives a generalized MWAT for
.
winter survival instead of the species
specific approach used in the other types of criteria.
All the lower incipient lethal temperature data from Appendix C for
freshwater fish species were used to calculate the regression line, which had
a slope of 0.50 and a correlation coefficient of 0.75. This regression line
was then displaced by approximately 2.5° C since it passed through the middle
. of the data and did not represent the more sensitive species. This new line
on the edge of the data array was then displaced by a 2° C safety factor, the
same factor discussed earlier, to account for the fact that the original data
points were for 50 percent survival and the 2° C safety factor would result
in 100 percent survival, These two adjustments in the original regression
line therefore result in a line (Figure 11 that should insure no more than
negligible mortality of any fish species, At lower acclimation temperatures
the coldwater species were different from the warmwater species, and the resulta
criterion takes this into account,
If fish can congregate in an area close to-the discharge point, this
criterion could be a limit on the degree rise permissible at a particular site.
Obviously, if there is a screened discharge channel in which some cooling
occurs, a higher initial discharge temperature could be permissible to fish.
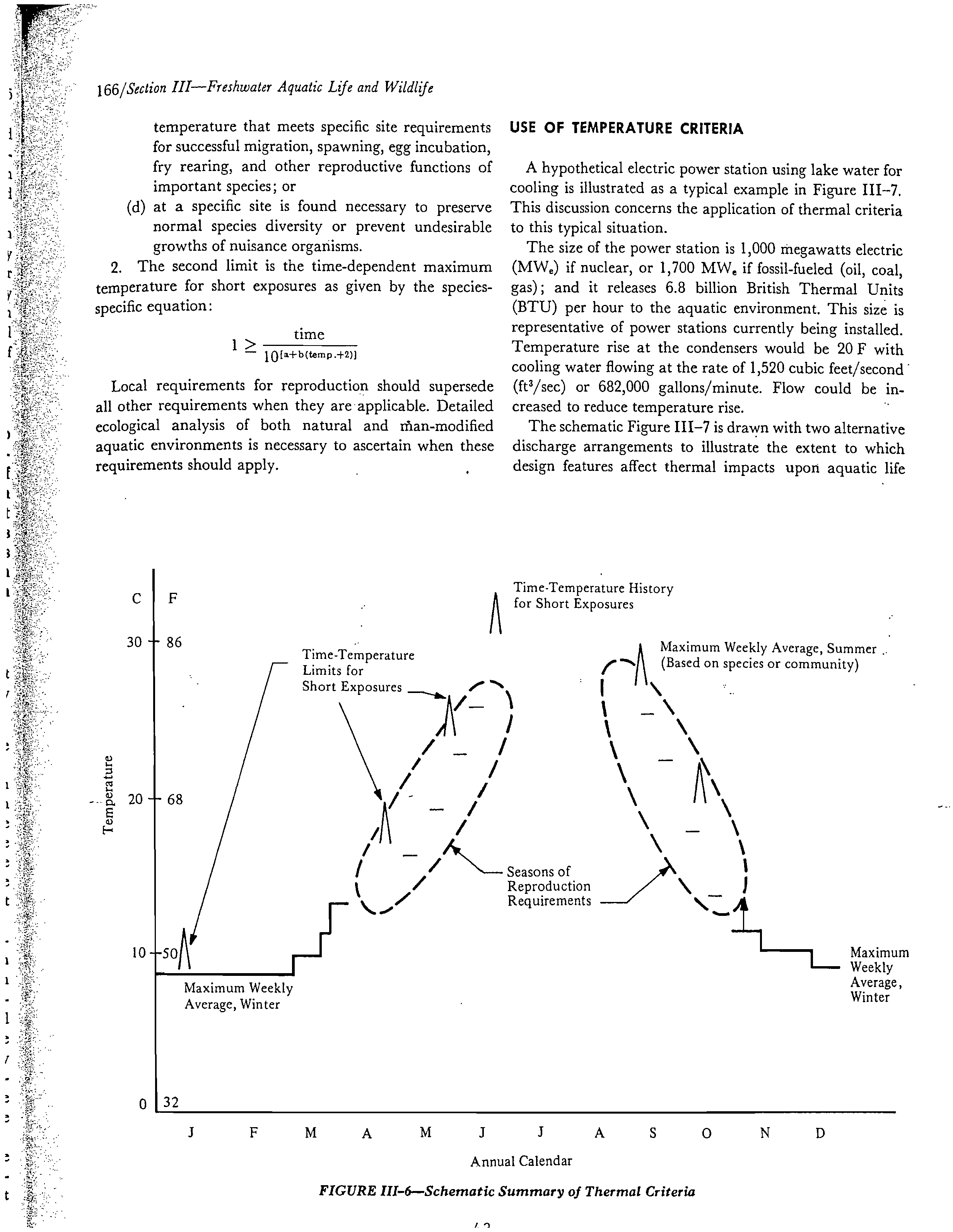
An example of the use of this criterion (as plotted in the nomograph,
Figure 1) would be a situation in which the ambient water temperature is 10°
C, and the MWAT, where fish could congregate, is 25° C, a difference of 15°
C. At a lower ambient temperature of about 2.5° C, the MWAT would be 10° C,
-a 7.5° C difference.
18
C:
30(86)
CC
< 25(77)
CC
LU
a
_
2
20(68)
2
a.
15(59)
—J
2
10(50)
CC
6(41)
•
0
sZ%
4./
,■.,
. P.—
WARMWATER
FISH SPECIES'
1
/
/C......... COLDWATER
/?
FISH SPECIES
/
../
I
5(41)
?
10(50)
AMBIENT TEMPERATURE
15(59)
Figure 1. Nomograph to determine the maximum weekly average
temperature of plumes for various ambient temperatures,
°C (°F).
SECTION 5
EXAMPLES
Again, because precise thermal-effects data are not available for all
species,• we would like to emphasize the necessity for subjective decisions
based on common-sense knowledge of existing aquatic systems. For some
fish species for which few or only relatively poor data are available,
subjectivity becomes important. If several qualified people were to calcula
various temperature criteria for species for which several sets of high qual
data were available, it is unlikely that they would be in agreement in all
instances.
The following examples for warmwater and coldwater species are presente
only as examples and are not at all intended to be water-body-specific
recommendations. Local extenuating circumstances may warrant differences,
the basic conditions of the examples may be slightly unrealistic. More
precise estimates of principal spawning and growth seasons should be
available from the local state fish departments.
EXAMPLE 1
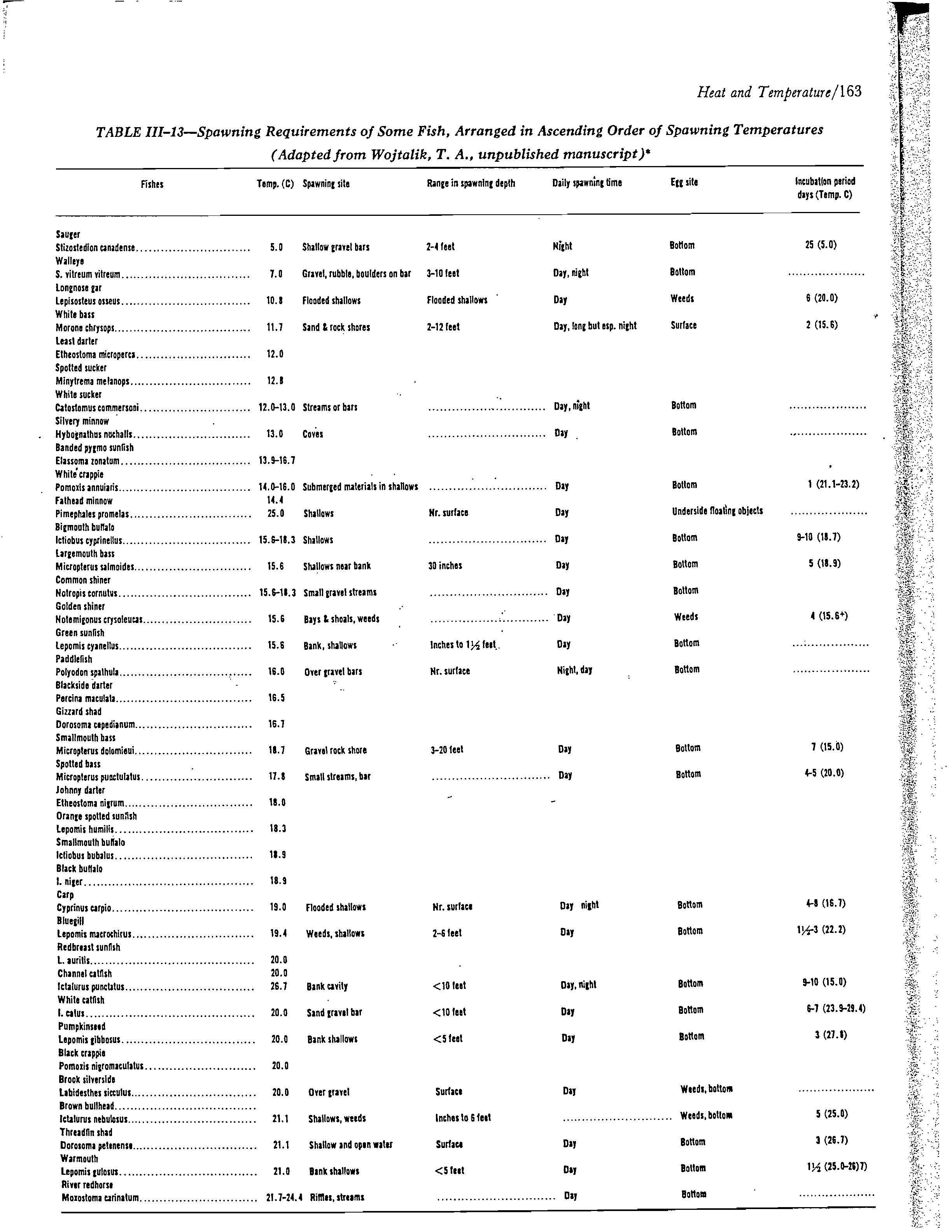
Tables 1 and 2, Figure 1, and Appendix C are the principal data sources
for the criteria derived for this example. The following water-body-specifi
data are necessary and in this example are hypothetical:
1.
Species to be protected by the criteria: channel catfish, largemc
bass, bluegill, white crappie, freshwater drum, and bigmouth buffalo.
2.
Local spawning seasons for these species: April to June for the
white crappie and the bigmouth buffalo; other species, May to July.
3.
Normal ambient winter temperature: 5° C
in December and January;
10° C in November, February, and March.
4.
The principal growing season for these fish species: July througt
September.
5.
Any local extenuating circumstances should be incorporated into tt
criteria as appropriate. Some examples would be yellow perch gamete
maturation in the winter, very temperature-sensitive endangered species,
or important fish-food organisms that are very temperature sensitive. For
the example we will have no extenuating circumstances.
20
In some instances the data will be insufficient to determine each
necessary criterion for each species. Estimates must be made based on
available species-specific data or by extrapolation from data for species with
similar requirements for which adequate data are available. For instance, this
example includes the bigmouth buffalo and freshwater drum for which no growth
or short-term summer maxima are available (Table 1). One would of necessity
have to estimate that the summer criteria would not be lower than that for the
white crappie, which has a spawning requirement as low as the other two
species.
The choice of important fish species is very critical. Since in this
example the white crappie is as temperature sensitive as any of the species,
the maximum weekly average temperature for summer growth is based on the
white crappie. Consequently, this criterion would result in lower than
optimal conditions for the channel catfish, bluegill, and largemouth bass.
An alternate approach would be to develop criteria for the single most
important species even if the most sensitive is not well protected. The
choice is
. a socioeconomic one.
Before developing a set of criteria such as those in Table 3, the material
material in Tables 1 and 2 should be studied for the species
of concern. It is
evident that the lowest optimum temperature for summer growth for the species
for which data are available would be for the white crappie (28° C). However,
there is no maximum for short exposure since the data are not available (Appendix,
C). For the species for which there are data, the lowest maximum for short
exposure is fon:the largemouth bass-(34° C). In this example we have all
the necessary data for spawning and maximum for short exposure for embryo
survival for all species of concern (Table 2).
During the winter, criteria may be necessary both for the mixing zone as
well as for the receiving water. Receiving-water criteria would be necessary
if an important fish species were known to have gamete-maturation requirements
like the yellow perch, or embryo-incubation requirements like trout, salmon,
cisco, etc, In this example there is no need for receiving-_system water criteria.
At this point, we are ready to complete Table 3 for Example 1.
EXAMPLE 2
All of the general concerns and data sources presented throughout the
discussion and derivation of Example 1 will apply here.
1. Species to be protected by the criteria: rainbow and brown trout
and the coho salmon.
2.
Local spawning seasons for these species: November through January
for rainbow trout; and November through December
for the brown trout and coho
salmon.
3.. Normal ambient winter temperature: 2° C in November through February;
5° C in October, March, and April.
21
TABLE 3.?
TEMPERATURE CRITERIA FOR EXAMPLE 1
Maximum weekly average temperature,
,
•
C (° F))
Month
Receiving water
Heated plume
Decision basis
January
__ a
15(59)
Figure 1
February
--a
25(77)
Figure 1
March
25(77)
Figure 1
April
18(64)b
White crappie spawning
May
21(70)
Largemouth bass spawning
June
25(77)
Bluegill spawning and
white crappie growth
July
28(82)
White crappie growth
August
28(82)
--
White crappie growth
September
28(82)
White crappie growth
October
iovember
21(70)
a
25(77)
Normal gradual seasonal
decline
Figure 1
December
__a
15(59)
Figure 1
Month
Short—term maximum
?
Decision basis
January
?
None needed
?
Control by MWAT in plume
February
?
None needed
?
Control by MWAT in plume
March
?
Nona needed
?
Control by MWAT in plume
April?
26(79)?
Largemouth bassb
survival
(estimated)
May
?
29(84)
?
Largemouth bass
b survival
(estimated)
June
?
34(93)
?
Largemouth bass' survival
July?
34(93)
?
Largemouth bassb
survival
August
?
34(93)
?
Largemouth bass
b survival
September?
34(93)
?
Largemouth basa
l
' survival
October
?
29(84)?
Largemouth basa
l' survival
(estimated) .
November?
None needed?
Control by MWAT in plume
December
?
None needed?
Control by MWAT in plume
a
If a species had required a winter chill period for gamete maturation'or egg incubation,
receiving—water criteria would also be required.
b
No data available for the'slightly more sensitive white crappie.
22
4.
The principal growing season for these fish species; June through
September.
5.
Consider any local extenuating circumstances: There are none in
this example.
At this point, we are ready to complete Table 4 for Example 2.
23
TABLE 4.
?
TEMPERATURE CRITERIA FOR EXAMPLE 2
Month
Maximum weekly average temperature,?
(' C (" F))
Receiving water?
Heated plume
Decision basis
January
9(48)
10(50)
Rainbow trout spawning
and Figure 1
February
13(55)
10(50).
Normal gradual seasonal
rise and Figure 1
March
13(55)
15(59)
Normal gradual seasonal
rise and Figure 1
April
14(57)
15(59)
Normal gradual seasonal
rise and Figure
1
May
16(61)
Normal gradual seasonal
rise
June
17(63)
Brown trout growth
July
17(63)
Brown trout growth
August
17(63)
Brown trout growth
September
17(63)
Brown trout growth
October
12(54)
15(59)
Normal gradual seasonal
decline
November
8(46)
10(50)
Brook trout spawning and
Figure 1
December
8(46)
10(50)
Brown trout spawning and
Figure 1
Month?
Short-term maximum
?
Decision basis
January 13(55) Embryo survival for
rainbow trout and
coho salmon
February 13(55) Embryo survival for
rainbow trout and
coho salmon
March 13(55) Embryo survival for
rainbow trout and
coho salmon
April
May
June
?
24(75)?
Short-term maximum for
survival of all species
July
?
24(75)
?
Short-term maximum for
survival of all species
August?
24(75)?
Short-term maximum for
survival of all species
September?
24(75)
?
Short-term maximum for
survival of all species -
October
November 13(55) Embryo survival for
rainbow trout and
coho salmon
December 13(55) Embryo survival for
rainbow trout and
coho salmon
24
REFERENCES
Brett, J. R. 1952. Temperature tolerance in young Pacific salmon, genus
Oncorhynchus. J. Fish, Res. Board Can. 9:265-323,
. 1956. Some principles in the thermal requirements of fishes.
Quart. Rev. Biol, 31:75-87.
Federal Water Pollution Control Administration. National Technical Advisory
Committee. 1968. Water Quality Criteria. U.S. Department of the Interior,
Washington, D.C. 245 p.
Federal Water Pollution Control Administration. 1969a. FWPCA Presentations
ORSANCO Engineering Committee. U.S. Department of the Interior,
Sixty-Ninth Meeting, Cincinnati, Ohio (May 13-14, 19691.
. 1969b. FWPCA Presentations ORSANCO Engineering Committee.
U.S. Department of the Interior, Seventieth Meeting, Cincinnati, Ohio
(September 10, 19691.
Fry, F. E. J., J, R, Brett, arid G. 11, Clawson. 1942. Lethal limits of
temperature for young goldfish. Rev. Can, Biol. 1:50-56,
Fry, F. E. J., J. S. Hart, and K, F. Walker. 1946. Lethal temperature
relations for a sample of young speckled trout, Salvelinus fontinalis.
Ontario Fish. Res. Lab, Pub, No. 66. Univ. Toronto Press, Toronto,
Can. pp. 9-35.
Great Lakes Water Quality Agreement. 1972. With Annexes and Texts and
Terms of Reference, Between the United States of America and Canada.
TS 548;36Stat.2448. (April 15, 1972
1 . 69 p.
McKee, J; E., and H. W. Wolf, 1963, Water Quality Criteria 12nd ed.].
The Resources Agency of California Pub. No. 3-A., State Water Quality
Control Board, Sacramento, Calif. 548 p.
National Academy of Sciences and National Academy of Engineering (NAS/NAE).
1973. Water Quality Criteria 1972. A Report of the Committee on Water
Quality Criteria. U.S. Environmental Protection Agency Pub. No. EPA-R3-
73-033. Washington, D.C. 553 p.
Ohio River Valley Water Sanitation Commission (ORSANC0). Aquatic Life Advisory
Committee. 1956. Aquatic life water quality criteria ---second progress
report. Sew. Ind. Wastes 28:678-690.
25
. 1967, Aquatic life water quality criteria /---fourth. progress
report, Env. Sci. Tech. 1:888-897.
?
. 1970. Notice of requirements (standards number 1-70 and 2-70)
pertaining to sewage and industrial wastes discharged to the Ohio
River. ORSANCO, Cincinnati, Ohio.
Public Law 92-500. 1472. An Act to Amend the Federal Water Pollution
Control Act. 92nd Congress, S. 2770, October 18, 1972. 86 STAT. 816
through 86 STAT 904.
U.S. Environmental Protection Agency. 1976. Quality Criteria for Water.
Office of Water and Hazardous Materials, Washington, D.C. EPA 440/9-
76-023, 501 p.
26
APPENDICES
Page
A?
Heat. and Temperature (from the National Academy of Sciences
and National Academy of Engineering, 1973) ?
28
B
?
Thermal Tables (from the National Academy of Sciences and
'?
National Academy of Engineering, 1973).
?
51'
C?
Fish Temperature'Data (° C)
?
62
27
APPENDIX A*
HEAT AND TEMPERATURE
Living organisms do not respond to the quantity of heat
but to degrees of temperature or to temperature changes
caused by transfer of heat. The importance of temperature
to
acquatic organisms is well known
. , and the composition
of aquatic communities depends largely on the temperature
characteristics of their environment. Organisms have upper
and lower thermal tolerance limits, optimum temperatures
for growth, preferred temperatures in thermal gradients,
and temperature limitations for migration, spawning, and
egg incubation. Temperature also affects the physical
environment of the aquatic medium, (e.g., viscosity, degree
of ice cover, and oxygen capacity. Therefore, the com-
position of aquatic communities depends largely on tem-
perature characteristics of the environment. In recent
years there has been an accelerated demand for cooling
waters for power stations that release large quantities of
heat, causing, or 'threatening to cause, either a warming of
rivers, lakes, and coastal waters, or a rapid cooling when the
artificial sources of heat are abruptly terminated. For these
reasons, the environmental consequences of temperature
changes must be considered in assessments of water quality
requirements of aquatic organisms.
The •"natural" temperatures of surface waters of the
United States vary from 0 C to over 40 C as a function of
latitude, altitude, season, time of day, duration of flow,
depth, and many other variables. The agents that affect
the natural temperature are so numerous that it is unlikely
that two bodies of water, even in the same latitude, would
have exactly the same thermal characteristics. Moreover, a
single aquatic habitat typically does not have uniform or
consistent thermal characteristics. Since all aquatic or-
ganisms (with the exception of aquatic mammals and a
few large, fast-swimming fish) have body temperatures that
conform to the water temperature, these natural variations
create conditions that are optimum at times, but are
generally above or below optima for• particular physio-
logical, behavioral, and competitive functions of the species
present.
Because significant temperature changes may affect the
composition of an aquatic or wildlife community, an
induced change in the thermal characteristics of an eco-
system may be detrimental. On the other hand, altered
thermal characteristics may be beneficial, .as evidenced
in
most fish hatchery practices and at other aquacultural
facilities. (See the discussion of Aquaculture in Section IV.)
The general difficulty in developing suitable criteria for
temperature (which would limit the addition of heat) lies
in determining the deviation from "natural" temperature a
particular body of water can experience without suffering
adverse effects on its biota. Whatever requirements are
suggested, a "natural" seasonal cycle must be retained,
annual spring and fall changes in temperature must be
gradual, and large unnatural day-to-day fluctuations
should be avoided. In view of the many variables, it seems
obvious that no single temperature requirement can be
applied uniformly to continental or large regional areas;
the requirements must be closely related to each body of
water and to its
-
particular community of organisms,
especially the important species found in it. These should
include invertebrates, plankton, or other plant and animal
life that may be of importance to food chains or otherwise
interact with species of direct interest toman. Since thermal
requirements of various species differ, the social choice of
the species to be protected allows for different
-"levels of
protection" among water bodies as suggested by Doudoroff
and Shumway (1970)
272 for dissolved oxygen criteria. (See
Dissolved Oxygen, p. 131.) Although such decisions clearly
transcend the scientific judgments needed in establishing
thermal criteria for protecting selected species, biologists can
aid in making them. Some measures useful in assigning
levels of importance to species are: (1) high yield to com-
mercial or sport fisheries, (2) large biomass in the existing
ecosystem (if desirable), (3) important links in food chains
of other species judged important for other reasons, and
(4) "endangered" or unique status. If it is desirable to
attempt strict preservation of an existing ecosystem, the
most sensitive species or life stage may dictate the criteria
selected.
Criteria for making recommendations for water tem-
perature to protect desirable aquatic life cannot be simply a
maximum allowed change from "natural temperatures."
This is principally because a change of even one degree from
*From: National Academy of Sciences (1973), See pp. 151-171, 205-207.
111
152/Section III—Freshwater Aquatic Life and Wildlife
an ambient temperature has varying significance for an
organism, depending upon where the ambient level lies
within the tolerance range. In addition, historic tempera-
ture records or, alternatively, the existing ambient tempera-
ture prior to any thermal alterations by man are not always
reliable indicators of desirable conditions for aquatic
populations. Multiple developments of water resources also
change water temperatures both upward (e.g., upstream
power plants or shallow reservoirs) and downward (e.g.,
deepwater releases from large reservoirs), so that "ambient"
and "natural" are exceedingly difficult to define at a given
point over periods of several years.
Criteria for temperature should consider both the multiple
thermal requirements of aquatic species and requirements
for balanced communities. The number of distance require-
ments and the necessary values for each require periodic
reexamination as knowledge of thermal effects on aquatic
species and communities increases. Currently definable
requirements include:
•
maximum sustained temperatures that are con-
sistent with maintaining desirable levels of pro-
ductivity;
•
maximum levels of metabolic acclimation to warm
temperatures that will permit return to ambient
winter temperatures should artificial sources of
heat cease;
•
temperature limitations for survival of brief exposures
to temperature extrenies, both upper and lower;
• restricted temperature ranges for various stages of
reproduction, including (for fish) gonad growth and
gamete maturation, spawning migration, release of
gametes, development of the embryo, commence-
ment of independent feeding (and other activities)
by juveniles; and temperatures required for meta-
morphosis, emergence, and other activities of lower
forms;
•
thermal limits for diverse compositions of species of
aquatic communities, particularly where reduction
in diversity creates nuisance growths of certain
organisms, or where important food sources or
chains are altered ;
•
thermal requirements of downstream aquatic life
where upstream warming of a cold-water source will
adversely affect downstream temperature require-
ments.
Thermal criteria must also be formulated with knowledge
of how man alters temperatures, the hydrodynamics of the
c
hanges, and how the biota can reasonably be expected to
interact with the thermal regimes produced. It is not
su
fficient, for example, to define only the thermal criteria
for sustained production of a species in open waters, because
large numbers of organisms may also be exposed to thermal
c
hanges by being pumped through the condensers and
mixing zone of a power plant. Design engineers need
29
particularly to know the biological limitations to their
design options in such instances. Such considerations may
reveal nonthermal impacts of cooling processes that may
outweigh temperature effects, such as impingement of fish
upon intake screens, mechanical or chemical damage to
zooplankton in condensers, or effects of altered current
patterns on bottom fauna in a discharge area. The environ-
mental situations of aquatic organisms (e.g., where they
are, when they are there, in what numbers) must also be
understood. Thermal criteria for migratory species should
be applied to a certain area only when the species is actually
there. Although thermal effects of power stations are
currently of great interest, other less dramatic causes of
temperature change including deforestation, stream chan-
nelization, and impoundment of flowing water must be
recognized.
DEVELOPMENT OF CRITERIA
Thermal criteria necessary for the protection of species or
communities are discussed separately below. The order of
presentation of the different criteria does not imply priority
for any one body of water. The descriptions define preferred
methods and procedures for judging thermal requirements,
and generally do not give numerical values (except in
Appendix II–C). Specific values for all limitations would
require a biological handbook that is far beyond the scope
of this Section. The criteria may seem complex, but they
represent an extensively developed framework of knowledge
about biological responses. (A sample application of these
criteria begins on page 166, Use of Temperature Criteria.)
TERMINOLOGY DEFINED
Some basic thermal responses of aquatic organisms will
be referred to repeatedly and are defined and reviewed
briefly here. Effects of heat on organisms and aquatic
communities have been reviewed periodically (e.g., Bullock
1955,
259 Brett 1956;2 " Fry 1947,
276
1964,
276
1967;2
" Kinne
1970
29
9. Some effects have been analyzed in the context of
thermal modification by power plants (Parker and Krenkel
1969; 308
Krenkel and Parker 1969;
298
Cairns 1968;
261 Clark
1969;
263
and Coutant 1970c
269
). Bibliographic information
is available from Kennedy and Mihursky (1967),
294
Raney
and Menzel (1969), 313 and from annual reviews published
by the Water Pollution Control Federation (Coutant
1968,2"
1969,
266 1970a,262
197 1270).
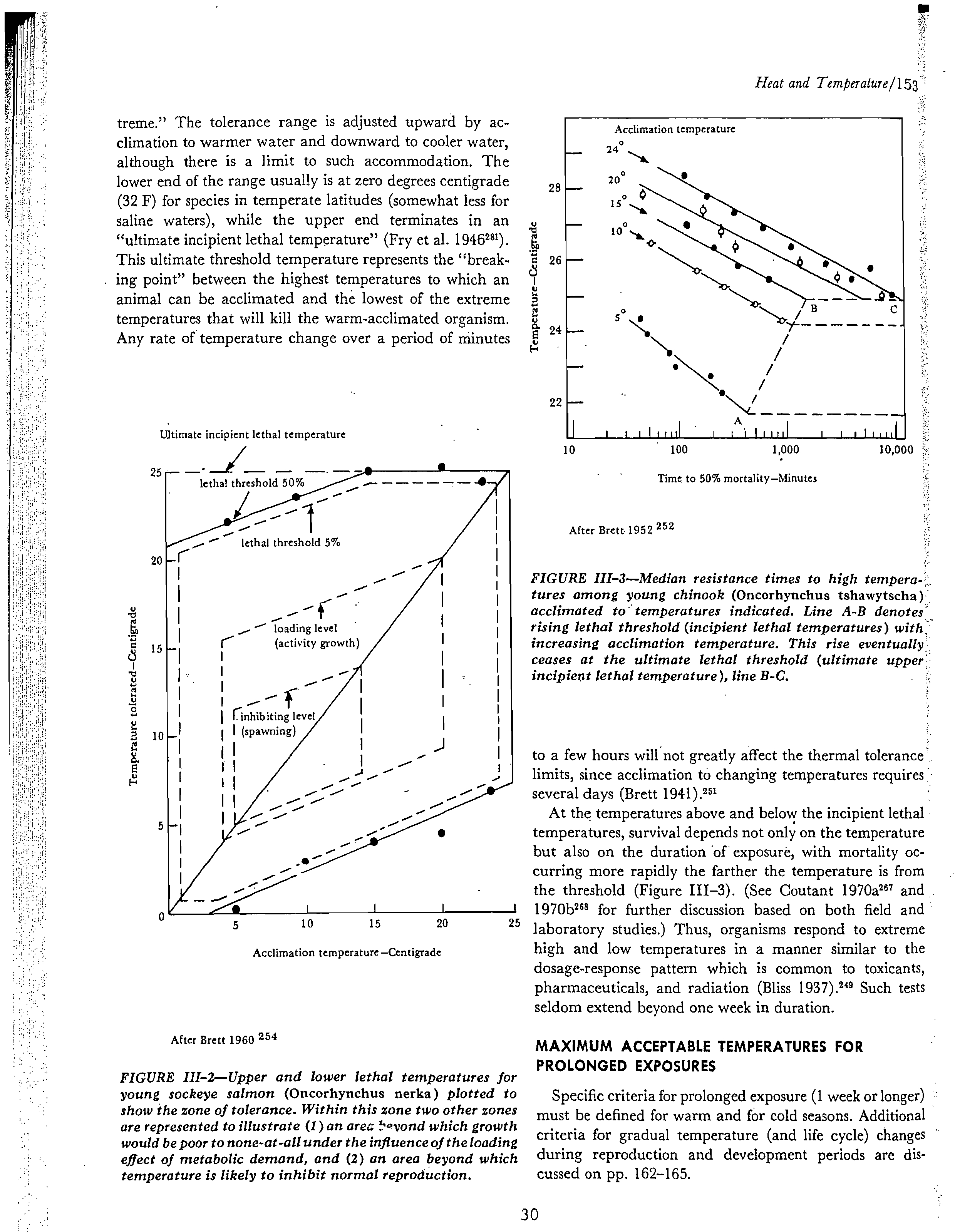
Each species (and often each distinct life-stage of a species)
has a characteristic tolerance range of temperature as a
consequence of acclimations (internal biochemical adjust-
ments) made while at previous holding temperature (Figure
111-2; Brett 1956
253
). Ordinarily, the ends of this range, or
the lethal thresholds, are defined by survival of 50 per cent
of a sample of individuals. Lethal thresholds typically are
referred to as "incipient lethal temperatures," and tem-
perature beyond these ranges would be considered "ex-
Ultimate incipient lethal temperature
25 —
lethal threshold 50%
lethal threshold 5%
r
?
loading
(activity
level
growth)
28
24
22
Heat and Temperature/153
treme." The tolerance range is adjusted upward by ac-
climation to warmer water and downward to cooler water,
although there is a limit to such accommodation. The
lower end of the range usually is at zero degrees centigrade
(32 F) for species in temperate latitudes (somewhat less for
saline waters), while the upper end terminates in an
"ultimate incipient lethal temperature" (Fry et al. 1946281).
This ultimate threshold temperature represents the "break-
ing point" between the highest temperatures to which an
animal can be acclimated and the lowest of the extreme
temperatures that will kill the warm-acclimated organism.
Any rate of temperature change over a period of minutes
5
Acclimation temperature—Centigrade
10?
100?1,000
Time to 50% mortality—Minutes
10,000
After Brett 1952
252
FIGURE III-3—Median resistance times to high tempera-
tures among
young
chinook
(Oncorhynchus tshawytscha)
acclimated to temperatures indicated. Line A-B denotes' _
rising lethal threshold (incipient lethal temperatures) with:
increasing acclimation temperature. This rise eventually
ceases at the ultimate lethal threshold (ultimate
upper
incipient lethal temperature), line B-C.
to a few hours will not greatly affect the thermal tolerance
limits, since acclimation to changing temperatures requires
several days (Brett 1941).2"
At the temperatures above and below the incipient lethal
temperatures, survival depends not only on the temperature
but also on the duration of exposure, with mortality oc-
curring more rapidly the farther the temperature is from
the threshold (Figure 111-3). (See Coutant 1970a
267 and
•
? I?
I
?
1970b2" for further discussion based on both field and
10?
15
?
20?
25
laboratory studies.) Thus, organisms respond to extreme
high and low temperatures in a manner similar to the
dosage-response pattern which is common to toxicants,
pharmaceuticals, and radiation (Bliss 1937). 2 " Such tests
seldom extend beyond one week in duration.
After Brett 1960 254
FIGURE III-2—Upper and lower lethal temperatures for
young
sockeye
salmon
(Oncorhynchus nerka)
plotted to
show the
zone
of tolerance. Within this zone two other zones
are represented to illustrate (1)
an
area !
•o
vond which growth
would be poor to none-at-all under the influence of the loading
effect of metabolic demand, and
(2) an
area beyond which
temperature is likely to inhibit normal reproduction.
MAXIMUM ACCEPTABLE TEMPERATURES FOR
PROLONGED EXPOSURES
Specific criteria for prolonged exposure (1 week or longer)
must be defined for warm and for cold seasons. Additional
criteria for gradual temperature (and life cycle) changes
during reproduction and development periods are dis-
cussed on pp. 162-165.
30
),0
00
pera-s.:
;cha)::
with
ually
ire
/
15
3
?
154 /Section 111—Freshwater Aquatic Life and Wildlife
SPRING, SUMMER, AND FALL MAXIMA FOR
PROLONGED EXPOSURE
ince
tires ..1'
thal
r7.
ture
OC-
70M
and
and
:me
the
nts,
ests
er
g
aal -
;es
fis-
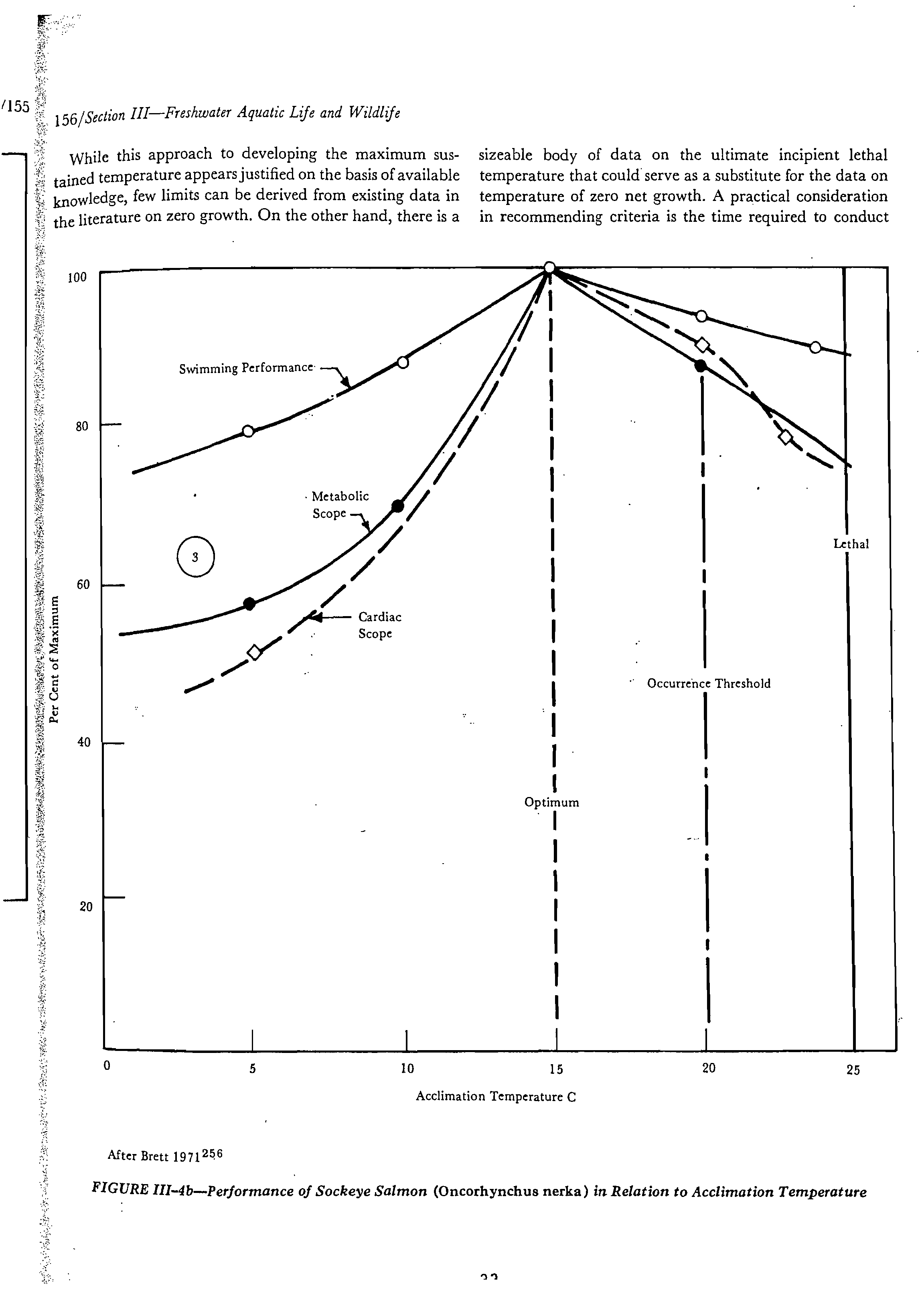
Occupancy of habitats by most aquatic organisms is
often limited within the thermal tolerance range to tem-
peratures somewhat below the ultimate upper incipient
lethal temperature. This is the result of poor physiological
performanc
e
at near lethal levels (e.g., growth, metabolic
scope for activities, appetite, _food conversion efficiency),
.interspecies competition, disease, predation, and other
subtle ecological factors (Fry 1951;
2
" Brett 1971
25
9. This
complex limitation is evidenced by restricted southern and
altitudinal distributions of many species. On the other hand,
optimum temperatures (such as those producing fastest
growth rates) are not generally necessary at all times to
maintain thriving populations and are often exceeded in
nature during 'summer months (Fry 1951;
2
" Cooper 1953;2"
Beyerle and Cooper 1960;
246
Kramer and Smith 1960207).
Moderate temperature fluctuations can generally • be
tolerated as long as a maximum upper limit is not exceeded
for long periods.
A true temperature limit for exposures long enough to
reflect metabolic acclimation and optimum ecological per-
formance must lie somewhere between the physiological
optimum and the ultimate upper incipient lethal tempera-
tures. Brett (1960)
2
" suggested that a provisional long-
term exposure limit be the temperature greater than opti-
mum that allowed 75 per cent of optimum performance.
His suggestion has not been tested by definitive studies.
Examination of literature on performance, metabolic
rate, temperature preference, growth, natural distribution,
and tolerance of several species has yielded an apparently
sound theoretical basis for estimating an upper temperature
limit for long term exposure and a method for doing this
with a minimum of additional research. New data will
provide refinement, but this method forms a useful guide
for the present time. The method is based on the general
observations summarized here and in Figure III-4(a, b, c).
1.
Performances of organisms over a range of tempera-
tures are available in the scientific literature for a variety of
functions. Figures III-4a and b show three characteristic
types of responses numbered 1 through 3, of which types 1
and 2 have coinciding optimum peaks. These optimum
temperatures are characteristic for a species (or life stage).
2.
Degrees of impairment from optimum levels of
various performance functions are not uniform with in-
creasing temperature above the optimum for a single species.
The most sensitive function appears to be growth rate, for
which a temperature of zero growth (with abundant food)
can be determined for important species and life stages.
Growth rate of organisms appears to be an integrator of all
factors acting on an organism. Growth rate should probably
be expressed as net biomass gain or net growth (McCormick
et al. 1971)
302
of the population, to account for deaths.
S. The maximum temperature at which several species
are consistently found in nature (Fry 1951;
2
" Narver
1970)
306
lies near the average of the optimum temperature
and the temperature of zero net growth.
4. Comparison of patterns in
.
Figures III-4a and b
among different species indicates that while the trends are
similar, the optimum is closer to the lethal level in some
species than it is in sockeye salmon. Invertebrates exhibit a
pattern of temperature effects on growth rate that is very
similar to that of fish (Figure III-4c).
The optimum temperature may be influenced by rate of
feeding. Brett et al. (1969)
257
demonstrated a shift in opti-
mum toward cooler temperatures for sockeye salmon when
ration was restricted. In a similar experiment with channel
catfish, Andrews and Stickney (1972)
2
" could see no such
shift. Lack of a general shift in optimum may be due to
compensating changes in activity of the fish (Fry
personal
observation). 326
These observations suggest that an average of the opti-
mum temperature and the temperature of zero net growth
[(opt. temp. + z.n.g. temp)/2J Would be a useful estimate of
a limiting weekly mean temperature for resident organisms,
providing the peak temperatures do not exceed values
recommended for short-term exposures. Optimum growth
rate would generally be reduced to no lower than 80 per cent
of the maximum if the limiting temperature is as 'averaged
above (Table III-11). This range of reduction from opti-
mum appears acceptable, although there are no quantita-
tive studies available that would allow the criterion to be
based upon a specific level of impairment.
The criteria for maximum upper temperature must allow
for seasonal changes, because different life stages of many
species will have different thermal requirements for the
average of their optimum and zero net growths. Thus a
juvenile fish in May will be likely to have a lower maximum
acceptable temperature than-will the same fish in July, and
this must be reflected in the thermal criteria for a waterbody.
TABLE 111-11—Summary of Some Upper Limiting
Temperatures
in C,
(f
or
periods longer
than one
week)
Based Upon
Optimum
Temperatures and Temperatures
of Zero Net Growth.
Species
Optimum?Zero net
growth
Reference
opt+z.n.g. % of
optimum
2
Catostomus commersoni (white sucker) ?21
29.6
•
28.3
86
Coregonus artedli (circa or lake herring).
? 16
21.2
McCormick et al.
18.6
82
1971"
Ictalurus punctatus (channel cash) ?30
35.7
Straws 1970
31
0
32.8
94
30
35.1
Andrews and Stickney
32.8
81
1972"
Lepomis macrochirus (bluegill) (year II)
?22
28.5
McComish 1971
501
25.3
82
Micropterus salmoides (largemouth bass) ?
21.5
34
Strewn 1951"
30.1
13
Notropis atherinoidss (emerald shiner)
?27
33
•
30.5
13
Saly
elinus tontinalis (brook trout) ?
15.4
18.8
•
17.1
10
*National Water Quality Laboratory, Duluth, Minn., unpublished data."
'1
1
100
Gross Conversion Efficiency,
90
Lethal
80
70
Digestion Rate
60
50
Occurrence Threshold
40
30
Growth Rate
Opt mum
20
10
Heat and Temperature/155
0
10
15
Acclimation Temperature C
20
?
25
After Brett 1971256
Maximum Meal Size
FIGURE III-la—Performance of Sockeye Salmon
(Oncorhynchus nerka) in
Relation
to
Acclimation Temperature
32
Swimming Performance
• Metabolic
Scope
Cardiac
Scope
/
I
°
Optimum
1
Lethal
Occurrence Threshold
156/Section III—Freshwater Aquatic Life and Wildlife
While this approach to developing the maximum sus-
tained temperature appears justified on the basis of available
knowledge, few limits can be derived from existing data in
the literature on zero growth. On the other hand, there is a
sizeable body of data on the ultimate incipient lethal
temperature that could serve as a substitute for the data on
temperature of zero net growth. A practical consideration
in recommending criteria is the time required to conduct
/155
100
80
60
20
0
5
10
15
20
25
Acclimation Temperature C
After Brett 1971256
FIGURE III-4b—Performance of Sockeye Salmon
(Oncorhynchus nerka) in
Relation to Acclimation Temperature
0
200
150 -
00
f33
et
•
00
0 0%
0
#
1
0.0
•
o
o
0.1?
0
•
0
fioo
0
?
e
90
er°
8
o?
0
0 0
c
0
g
?
• o
o
0 •
o
%.
•0 a•
O1
°
o
k arm iiiissi 1 i eel
9?
5
10
15
20
25
Temperature in C
0
o
0
50
0
Heat and Temperature/157
research necessary to provide missing data. Techniques for
determining incipient lethal temperatures are standardized
(Brett 1952)
2" whereas those for zero growth are not.
A temperature that is one-third of the range between the
optimum temperature and the ultimate incipient lethal
temperature that can be calculated by the formula
ultimate incipient lethal temp.-optimum temp.
optimum temp. + ?
3
(Equation 1)
yields values that are very close to (optimum temp. +
z.n.g. temp.)/2. For example, the values are, respectively,
32.7 and 32.8 C for channel catfish and 30.6 and 30.8 for
largemouth bass (data from Table 111-8 and Appendix
. II).
This formula offers a practical method for obtaining allow- .
Ansell 1968
243
FIGURE III-4c—M.
mercenaria:
The general relationship
between temperature and the rate of shell growth, based
on
field measurements of growth and temperature.
sites in Poole Harbor, England; 0: North American sites.
able limits, while retaining as its scientific basis the require-
ments of preserving adequate rates of growth. Some limits
obtained from data in the literature are given in Table
111-12. A hypothetical example of the effect of this limit on
growth of largemouth bass is illustrated in Figure 111-5.
Figure 111-5 shows a hypothetical example of the effects
of the limit on maximum weekly average temperature on
growth rates of juvenile largemouth, bass. Growth data as a
function of temperature are from Strawn 1961 319
; the ambi-
ent temperature is an averaged curve for Lake Norman,
N. C., adapted from data supplied by Duke Power Com-
pany. A general temperature elevation of 10 F is used to
provide an extreme example. Incremental growth rates
(mm/wk) are plotted on the main figure, while annual ac-
cumulated growth is plotted in the inset. Simplifying as- '
sumptions were that growth rates and the relationship of '
growth rate to temperature were constant throughout the
year, and that there would be sufficient food to sustain
maximum attainable growth rates at all times.
The criterion for a specific location would be determined
by the most sensitive life stage of, an important species
likely to be present in that location at that time. Since
many fishes have restricted habitats (e.g., specific depth
zones) at many life stages, the thermal criterion must be
applied to the proper zone. There is field evidence that fish
avoid localized areas of unfavorably, warm water. This has
been demonstrated both in lakes where Coldwater fish
normally evacuate warm shallows in summer (Smith
1964) 3 '8
and at power station mixing zones (Gammon
1970;2 " Merriman et al. 1965).
304
In most large bodies of
water there are both vertical and horizontal thermal
gradients -that mobile organisms can follow to avoid un-
favorable high (or low) temperatures.
The summer maxima need not, therefore, apply to
mixing zones that occupy a small percentage of the suitable
habitat or necessarily to all zones where organisms have
free egress to cooler water. The maxima must apply, how-
ever, to restricted local habitats, such as lake hypolimnia or
thermoclines, that provide important summer sanctuary
areas for cold-water species. Any avoidance of a warm area
not part of the normal seasonal habitat of the species will
mean that less area of the water body is available to support
the population and that production may be reduced. Such
reduction should not interfere with biological communities
or populations of important species to a degree that is
damaging to the ecosystem or other beneficial uses. Non-
mobile organisms that must remain in the warm zone will
probably be the limiting organisms for that location. Any
recommendation for upper limiting temperatures must be
applied carefully with understanding of the population
dynamics of the species in question in order to establish
both local and regional requirements.
Elevated (with limit)
Elevated (without limit)
Average Ambient.,
(Lake Norman, N.C.)
Weekly Growth Rate
(Ambient + 10 F)
Weekly Growth Rate
(Ambient)
Ambient + 10 F
158 I Section III—Freshwater Aquatic Life and Wildlife
Annual Accumulated Growth
250
6 200
-5
150
60
U
50
2?
4?
6?
8?
10?
12
Weeks
6
3
•
2 5
4
10
9
8
7
2
3
i?
I
?
I
?
I?
I?
I?
I
?
I?
I?
I?
I?
I?
I
?
i
?
l?
I?
I?
I?
I?
I?
I?
I?
I?
I
?0?
7 14 21?
28 4 11 18?
25 4?
11 18 25
?
1
?
8 15 22 29 6
?
13 20 27 3 10 17 24 1
JAN.
?
FEB.
?
MAR.?
APR.
?
MAY
?
JUNE
35
U
U
22
20
18
16
14
12
10
8
6
4
Heat and Temperature/ 159
FIGURE III-5—A hypothetical example of the effects of the limit
on maximum
weekly
average temperature on growth rates of juvenile
largemouth
bass. Growth
data
as a function
of temperature are from Strawn 1961; the ambient temperature is
an
averaged curve for Lake
Norman, N.C., adapted from data supplied by Duke Power
Company. A
general temperature
elevation of 10 F is used to provide
an
extreme example. Incremental growth rates (mm/wk)
are plotted on the main figure, while annual accumulated growth is plotted in the inset.
Simplifying assumptions were that growth rates and the relationship of growth rate to
tem-
perature
were constant throughout the year, and that there would be sufficient food to sus-
tain maximum attainable growth rates at all times.
'0— Max. Weekly Avg., largemouth bass
••••■•■1
111111111111111_1111111111
3
15
22
29
5 12 19 26 2
9 16 23 30
7 14 21 28 4
11
18 25 2
9 16 23 30
JULY
AUG.
SEPT.
OCT.
NOV.
DEC.
with limit
Incremental
Growth Rates
(mm/wk)
I
without limit /
/
38
36
34
32
30
28
26
24
Micropterus dolomitic (smallmouth bass)._
Micropterussalmoides (largemouth bass)(try).
21.5
Notropis atherinoides (emerald shiner) .....
21
Oncorhynchus nerka (sockeye salmon)
?
15.0
15.0
(juveniles)
?
15.0
Pseudopleuronectes Americanus (winter
flounder)
?
18.0
Sams
truth (brown trout)
?
Ito 11
ave 12.5
Salvelinus footinalis (brook trout)
?
Salvolinus namaycush (lake trout) ?
26.3
?
83?
growth
28.3?
83
?
• growth
ave 21.3?
81.1
11.5?
growth
80.6?
growth
59.0?
growth
59.0•
?
other functions
max. swimming
64.4?
growtt7
54.5?
growth
15.4
?
59.1
?
growth
13.0?
55.4
?
growth
15?
59?
metabolic
ave 14.5?
53.1?
scope
16?
60.8?
scope tor activity
(2 metabolism)
17?
62.6
?
swimming speed
ave 16.5?
61.1
160/Section III-Freshwater Aquatic Life and Wildlife
TABLE m-12-Summary of Some Upper Limiting Temperatures for Prolonged Exposures of Fishes Based on
Optimum Tem-
peratures
and Ultimate Upper Incipient Lethal Temperatures (Equation 1).
Reference
Ultimate upper incipient
lethal temperature
C
Reference
Maximum weekly average
temperature (Eq. 1)
C?
F
unpubl., NWC11.324
29.3
84.1
Hart 1941"
21.8
82
McCormick
et
a1.1971'02
25.1
71.3
Edsall and Colby 19102
2
4
19.2
66.6
Straws 1970;
420
Andrews and Stickney
38.0
100.4
Allen and Strewn 1961
140
32.1
90.9
1911242
McComish 1971"
33.8
92.8
Hart 19522
44
25.9
11.6
Anderson 1959241
Horning and Pearson 1972291
35.0
95.0
Horning and Pearson 1912
241
29.9
85.8
Peek 196520
Strewn 196149
36.4
91.5
Hera 1952446
30.5
86.7
unpubL, NWEI1.321
30.1
81.3
Hart 1952244
21.2
82.1
Brett et a1.1969257
25.0
71.0
Brett 1952242
18.3
64.9
Brett 1971244
Brett 1970244
29.1
84.4
HMI and Westman 1966"
21.1
11.2
Brett 1910245
23.5
74.3
Bishai 1960247
16.2
61.2
unpubl, NWOL42s
25.5
11.9
Fry, Hart
and
Walker, 1946241
11.2
64.8
Baldwin 1951244
Graham 1949444
Gibson and Fry 1954"
23.5
Gibson and Fry 1954
213
18.8
65.8
Optimum
Species
?
Function
C?
F
Catostomus commersoni (white sucker)
??
27?
80.6
?
growth
Coregonus artedli (Cisco or rake herring) •
?
16?
60.1?
growth
Ictaluros punctatus (channel catfish)
??
30
?
86?
growth
Lepomis macrochiros (bluegill) (yr 11)
??
22?
71.6?
growth
Heat added to upper reaches of some cold rivers can be
retained throughout the river's remaining length ( Jaske
and Synoground 1970).
292
This factor adds to the natural
trend of warming at distances from headwaters. Thermal
additions in headwaters, therefore, may contribute sub-
stantially to reduction of cold-water species in downstream
areas (Mount 1970).
3
" Upstream thermal additions should
be evaluated for their effects on summer maxima at down-
stream locations, as well as in the immediate vicinity of
the heat source.
Recommendation
Growth of aquatic
organisms would be main-
tained at levels necessary for sustaining actively
growing and reproducing populations if
the maxi-
mum weekly average temperature in the zone in-
habited by the species
at that time does not exceed
o
ne-third of the range between the optimum tem-
p
erature and the ultimate upper incipient lethal
te
mperature of the species (Equation
1,
page 157),
and the temperatures above the weekly average do
not exceed the criterion for
short-term exposures.
This maximum need
not apply to
acceptable mix-
ing zones (see proportional relationships of mixing
zones to receiving systems, p. 114), and must be
ap
se
asonal
plied with
distribution
adequate
of
understanding
the important
of
species.
the normal
WINTER MAXIMA
Although artificially produced temperature elevations
during winter months may actually bring the temperature
closer to optimum or preferred temperature for important
species and attract fish (Trembley 1965),
3
" metabolic
acclimation to these higher levels can preclude safe return
of the organism to ambient temperatures should the
artificial heating suddenly cease (Pennsylvania Fish Com-
mission 1971;
2
" Robinson 1970)
3
" or the organism be
driven from the heat area. For example, sockeye salmon
(Oncorhynchus nerka)
acclimated to 20 C suffered 50 percent
mortality in the laboratory when their temperature was
dropped suddenly to 5 C (Brett 1971:
2
" see Figure
111-3).
The same population of fish withstood a drop to zero when
acclimated to 5 C. The lower limit of the range of thermal
tolerance of important species must, therefore, be main-
tained at the normal seasonal ambient temperatures
throughout cold seasons, unless special provisions are made
to assure that rapid temperature drop will not occur or that
organisms cannot become acclimated to elevated tempera-
tures. This can be accomplished by limitations on tempera-
ture elevations in such areas as discharge canals and mixing
zones where organisms may reside, or by insuring that
maximum temperatures occur only in areas not accessible
to important aquatic life for lengths of time sufficient to
allow metabolic acclimation. Such inaccessible areas would
include the high-velocity zones of diffusers or screened
Heat and Temperature/ 161
charge channels. This reduction of maximum temperatures
would not preclude use of slightly warmed areas as sites for
intense winter fisheries.
This consideration may be important in some regions at
times other than in winter. The Great Lakes, for example,
are susceptible to rapid changes in elevation of the thermo-
cline in summer which may induce rapid decreases in
shoreline temperatures. Fish acclimated to exceptionally
high temperatures in discharge canals may be killed or
severely stressed without changes in power plant opera-
tions (Robinson 1968). 3 " Such regions should take special
note of this possibility.
Some numerical values for acclimation temperatures and
lower limits of tolerance ranges (lower incipient lethal
temperatures) are given in Appendix II–C. Other data must
be provided by further research. There are no adequate
data available with which to estimate a safety factor for no
stress from cold shocks. Experiments currently in progress,
however, suggest that channel catfish fingerlings are more
Susceptible to predation after being cooled more than 5 to
6 C (Coutant,
unpublished data).324
The effects of limiting ice formation in lakes and rivers
should be carefully observed. This aspect of maximum
winter temperatures is apparent, although there is insuffi-
cient evidence to estimate its importance.
Recommendation
Important species should be protected if the
maximum weekly average temperature during win-
ter months in any area to which they have access
does not exceed the acclimation temperature
(minus a 2 C safety factor) that raises the lower
lethal threshold temperature of such species above
the normal ambient water temperatures for that
season, and the criterion for short-term exposures
is not exceeded. This recommendation applies es-
pecially to locations where organisms may be at-
tracted from the receiving water and subjected to
rapid thermal drop, as in the low velocity areas of
water diversions (intake or discharge), canals, and
mixing zones.
SHORT-TERM EXPOSURE TO EXTREME TEMPERATURE
To protect aquatic life and yet allow other uses of the
water, it is essential to know the lengths of time organisms
can survive extreme temperatures (i.e., temperatures that
exceed the 7-day incipient lethal temperature). Both
natural environments and power plant cooling systems can
briefly reach temperature extremes (both upper and lower)
without apparent detrimental effect to the aquatic life
(Fry 1951;
2
" Becker et al. 1971).2"
The length of time that 50 per cent of a population will
ci■ruive terrinerature ahrwe the inrinient lethal ternnerature
can be calculated from a regression equation of experi.
mental data (such as those in Figure 111-3) as follows:
log (time) = a+ b (temp.)
(Equation 2)
where time is expressed in minutes, temperature in degrees,
centigrade and where a and b are intercept and slope;
respectively, which are characteristics of each acclimationl
temperature for each species. In some cases the time•1
temperature relationship is more complex than the semi.'
logarithmic model given above. Equation 2, however, is
the most applicable, and is generally accepted by thel
scientific community (Fry 1967).279
Caution is recom-
mended in
.
extrapolating beyond the data limits of thel
original research (Appendix II–C). The rate of temperature
change does not appear to alter this equation, as long as thel
change occurs more rapidly than over several days (Brett
1941;
251
Lemke 1970).
30
Thermal resistance may be'
diminished by the simultaneous presence of toxicants or:
other debilitating factors (Ebel et al. 1970,213
and summaryi
by Coutant 1970c). 2
" The most accurate predictability can
be derived from data collected using water from the site]
under evaluation.
Because the equations based on research on thermal
tolerance predict 50 per cent mortality, a safety factor
needed to assure no mortality. Several studies have indi-
c ted that a 2 C reduction of an upper stress temperature',
results in no mortalities within an equivalent exposure
duration (Fry et al. 1942;
280
Black 1953).
248
The validity`;
of a two degree safety factor was strengthened by the results
of Coutant (1970a).
267
He showed that about 15 to 20
per cent of the exposure time, for median mortality at a given
high temperature, induced selective predation on thermally;
shocked salmon and trout. (This also amounted to reduction
of the effective stress temperature by about 2 C.) Un- ,
published data from subsequent predation experiments
showed that this reduction of about 2 C also applied to the
incipient lethal temperature. The level at which there is no
increased vulnerability to predation is the best estimate of a
no-stress exposure that is currently available. No similar
safety factor has been explored for tolerance of low tem-
peratures. Further research may determine that safety
factors, as well as tolerance limits, have to be decided
independently for each species, life stage, and water quality
situation.
Information needed for predicting survival of a number
of species of fish and invertebrates under short-term condi-
tions of heat extremes is presented in Appendix II–C. This
information includes (for each acclimation temperature)
upper and lower incipient lethal temperatures: coefficients
a and b for the thermal resistance equation ; and information
on size, life stage, and geographic source of the species.
It is clear that adequate data are available for only a small
percentage of aquatic species, and additional research is
necessary. Thermal resistance information should be
time
1 > 10
(a+b(temP •"+"2))
(Equation 3)
is
Le:
ie
Ie
)r
•y
:e
.t'
is
-e
:y
l y
a-
??s,"
ts
50
a
in
ar
ty
t.d
ty
er.
li-
iis
its
an
es.
all
is
be
al-
1621 Section III—Freshwater Aquatic Life and Wildlife
taneous presence of toxicants or other debilitating factors,
a consideration not reflected in Appendix II–C data. More
data are available for upper lethal temperatures than for
lower.
The resistance time equation, Equation 2, can be
rearrange
d
to incorporate the 2 C margin of safety and also
to define conditions for survival (right side of the equation
less than or equal to 1) as follows:
Low levels of mortality of some aquatic organisms are not
necessarily detrimental to ecosystems, because permissible
mortality levels can be established. This is how fishing or
shellfishing activities are managed. Many states and inter-
national agencies have established elaborate systems for
setting an allowable rate of mortality (for sport and com-
mercial fish) in order to assure needed reproduction and
survival. (This should not imply, however, that a form of
pollution should be allowed to take the entire harvestable
yield.) Warm discharge water from a power plant may
sufficiently stimulate reproduction of some organisms (e.g.,
zooplankton), such that those killed during passage through
the maximally heated areas are replaced within a few hours,
and no impact of the mortalities can be found in the open
water (Churchill and Wojtalik 1969;
2
" Heinle 1969).288
On the other hand, Jensen (1971)
293 calculated that even
• five percent additional mortality of 0-age brook trout
(Salvelinus fontinalis)
decreased the yield of the trout fishery,
and 50 per cent additional mortality would, theoretically.
cause extinction of the population. Obviously, there can be
no adequate generalization concerning the impact of short-
term effects on entire ecosystems, for each case will be
somewhat different. Future research must be directed
toward determining the effects of local temperature stresses
on population dynamics. A complete discussion will not be
attempted here. Criteria for complete short-term protection
may not always be necessary and should be applied with an
adequate understanding of local conditions.
Recommendation
Unless there is justifiable reason to believe it
unnecessary for maintenance of populations of a
species, the right side of Equation 3 for that
Species should not be allowed to increase above
unity when the temperature exceeds the incipient
lethal temperature minus 2 C:
1 >
— 10
(a+b(temp.+2))
Values for
a
and
b
at the appropriate acclimation
t
emperature for some species can be obtained from
Appendix II—C or through additional research if
necessary data are not available. This recommen-
dation applies to all locations where organisms to
be protected are exposed, including areas within
mixing zones and water diversions such as power
station cooling water.
REPRODUCTION AND DEVELOPMENT
The sequence of events relating to gonad growth and
gamete maturation, spawning migration, release of gametes,
development of the egg and embryo, and commencement
of independent feeding represents one of the most complex
phenomena in nature, both for fish (Brett 1970)
255
and
invertebrates (Kinne 1970).296
These events are generally
the most thermally sensitive of all life stages. Other environ-
mental factors, such as light and salinity, often seasonal in
nature, can also profoundly affect the response to tempera-
ture (Wiebe 1968).
323
The general physiological state of the
organisms (e.g., energy reserves), which is an integration of
previous history, has a strong effect on reproductive poten-
tial (Kinne 1970).2
" The erratic sequence of failures and
successes of different year classes of lake fish attests to the
unreliability of natural conditions for providing optimum
reproduction.
Abnormal, short-term temperature fluctuations appear to
be of greatest significance in reduced production of juvenile
fish and invertebrates (Kinne, 1963).
2
" Such thermal
fluctuations can be a prominent consequence of water use
as in hydroelectric power (rapid changes in river flow rates),
thermal electric power (thermal discharges at fluctuating
power levels), navigation (irregular lock releases), and
irrigation (irregular water diversions and wasteway re-
leases). Jaske and Synoground (1970)
292
have documented
such temperature changes due to interacting thermal and
hydroelectric discharges on the Columbia River.
Tolerable limits or variations of temperature change
throughout development, and particularly at the most
sensitive life stages, differ among species. There is no
adequate summary of data on such thermal requirements
for successful reproduction. The data are scattered through
many years of natural history observations (however, see
Breder and Rosen 1966
250
for a recent compilation of some
data ; also see Table III-13). High priority must be assigned
to summarizing existing information and obtaining that
which is lacking.
Uniform elevations of temperature by a few degrees
during the spawning period, while maintaining short-term
temperature cycles and seasonal thermal patterns, appear
to have little overall effect on the reproductive cycle of
resident aquatic species, other than to advance the timing
for spring spawners or delay it for fall spawners. Such shifts
are often seen in nature, although no quantitative measure-
ments of reproductive success have been made in this
connection. For example, thriving populations of many
fishes occur in diverse streams of the Tennessee Valley in
which the date of the spawning temperature may vary in a
time
Heat and Temperature/163
TABLE 111-13-Spawning Requirements of Some Fish, Arranged
in
Ascending Order of Spawning Temperatures
(Adapted from Wojtalik, T. A., unpublished manuscript)•
Fishes?
Temp. (C) Spawning site?
Range in spawning depth?
Daily spawning time?
Egg site
?
Incubation period
days (Temp. C)
Sauger
Stizostedion anadense
?
?
5.0?
Shallow gravel bars?
2-4 feet?
Night?
Bottom?
25 (5.0)
Walleye
S. vitreum vitreum ?
?
1.0?
Gravel, rubble, boulders on bar?
3-10 feet?
Day, night
?
Bottom
Longnose gar
Lepisosteus omen ?
?
10.8?
Flooded shallows?
Flooded shallows .
?
Day?
Weeds?
6 (20.0)
White bass
Morons chrysops ?
?
11.1?
Sand t rock shores?
2-12 feet?
Day, tong but esp. night?
Surface?
2 (15.6)
Least darter
Etheostoma microperca
?
?
12.0
Spotted sucker
Minytrema me lanops
?
?
12.8
White sucker
Catostomus commersoni ?
12.0-13.0 Streams or bars
?
?
Day, night
?
Bottom
Silvery minnow
Hybognathus =halls ?
?
13.0?
Coves?
?
Day .?
Bottom
Banded pygmo sunfish
Elassoma sonatom
?
13.9-16.7
White crappie
Pomovis annuiaris ?
14.0-16.0 Submerged materials in shallows
?
Day
?
Bottom
?
1 (21.1-23.2)
Fathead minnow?
14.4
Pi mephales promelas
?
?
25.0?
Shallows?
Hr. surface
?
Day
?
Underside floating objects
Bigmouth buffalo
Ictiobus cyminellus
?
15.6-18.3 Shallows?
?
Day
?
Bottom
?
9-10 (18.7)
Largemouth bass
Micropterus salmoides ?
?
15.6?
Shallows near bank
?30 inches
?
Day?
Bottom
?
5 (18.9)
Common shiner
flattops ornutus
?
15.6-11.3 Small gravel streams
?
?
Day
?
Bottom
Golden shiner
Notemigonus crysoleucas ?
?
15.6
?
Bays I shoals, weeds
?
?
Day?
Weeds
?
4 (15.6+)
Green sunfish
Lepomis cyanellus ?
?
15.6?
Bank, shallows
?
Inches to 1% feet.
?
Day?
Bottom
Paddlefish
Polyodon spalhula ?
?
16.0
?
Over gravel bars
?
Hr. surface
?
Night, day
?
Bottom
Blackside darter
?
-
Pemba maculata ?
?
16.5
Gizzard shad
Dorosoma rapedianum
?
?
16.7
Smallmouth bass
Micropterus dolomieui ?
?
18.7?
Gravel rock shore
?3-20 tent
?
Day?
Bottom
?
1 (15.0)
Spotted bass
Micropterus puottufalus ?
?
11.8?
Small streams, bar
?
?
Day
?
Bottom
?
4-5 (20.0)
Johnny darter
Etheostoma nivum ?
?
18.0
?
-
Orange spotted sunfish
Lepomis humilis ?
?
18.3
Smallmouth buffalo
Ictiobus bubalus ?
?
11.9
Black buffalo
I. niger
?
?
18.9
Carp
Cyprinus carpio ?
?
19.0
?
Flooded shallows
?Hr. surface
?
Day night
?
Bottom
?
4-8 (16.1)
Bluegill
Lepomis macrochirus
?
?
19.4?
Weeds, shallows
?2-6 feet
?
Day?
Bottom
?
114-3 (22.2)
Redbreast sunfish
L. auritis ?
?
20.0
Channel catfish?
20.0
Ictalurus punctatus ?
?
26.7?
Bank cavity
?
<10 feet
?
Day, night
?
Bottom
?
9-10 (15.0)
White catfish
I. catus ?
?
20.0
?
Sand gravel bar
?
<10 feet
?
Day?
Bottom
?
6-1 (23.9-29.4)
Pumpkinseed
Lepomis glbbosus.
?
?
20.0?
Bank shallows
?
<5 feet
?
Day?
Bottom
?
3 (21.1)
Black crappie
Pomoxis nigromaculatut .
?
20.0
Brook silverside
Labidesthes sicculus ?
?
20.0?
Over gravel?
Surface?
Day?
Weeds, bottom
Brown bullhead ?
Ictalurus nebulous
?
?
21.1?
Shallows, weeds
?
Inches to 6 feet
?
?
Weeds, bottom
?
5 (25.0)
Threadfin shad
Dorosoma petenense ?
?
21.1?
Shallow and open water?
Surface?
Day?
Bottom
?
3 (26.7)
Warmouth
Lepomis
1111011.11 ?
?
21.0
?
Bank shallows
?
<5 test
?
Day?
Bottom
?
114 (25.0-26)7)
River redhorse
Mosostoma urinatum ?
21.1-24.4 Riffles, streams?
?
Day
?
Bottom
164/Section
III—Freshwater Aquatic Life and Wildlife
TABLE III-13—Spawnin
g
Requirements of Some Fish, Arranged in Ascending Order of Spawning Temperatures—Continued
Fishes?
Temp. (C)?
Spawning site
?
Range in spawning depth
?
Daily spawning time?
Egg site?
Incubation period
days (Temp. C)
. Blue catfish
Ictalurus turcatus
?
?
22.2
Flathead catfish
Pylodictis oli
y
aris
?
?
22.2
&OW
sunfish
Lepomis microlophus
?
?
23.0?
Quiet, various
?Inches to 10 feet
1.0111e3f
sunfish
L megatons
?
?
23.3
Freshwater drum
Apiodinotus grunniens
?
?
23.0
River
carpsucker
Carpoides carps
?
?
23.9
Spotted bullhead
!dater serracanthus
?
?
26.7
Yellow bullhead
I. WM
?
?
Quiet, shallows
?1M-1 feet
Bottom?
5-10
(18.9)
* T. A.
Wojtalik, Tennessee Valley Authority, Muscle Shoals, Alabama.zn
given year by 22 to 65 days. Examination of the literature
shows that shifts in spawning dates by nearly one month
are common in natural waters throughout the U.S. Popula-
tions of some species at the southern limits of their dis-
tribution are exceptions, e.g., the lake whitefish
(Coregonus
clupeaformis)
in Lake Erie that require a prolonged, cold
incubation period (Lawler 1965) 299
and species such as
yellow perch
(Percaflavescens)
that require a long chill period
for egg maturation prior to spawning ( Jones,
unpublished
data).3"
This biological plasticity suggests that the annual spring
rise, or fall drop, in temperature might safely be advanced
(or delayed) by nearly one month in many regions, as long
as the thermal requirements that are necessary for migra-
tion, spawning, and other activities are not eliminated and
the necessary chill periods, maturation times, or incubation
periods are preserved for important species. Production of
food organisms may advance in a similar way, with little
disruption of food chains, although there is little evidence to
support this assumption (but see Coutant 1968;
2
" Coutant
and Steele 1968;
271
and Nebeker 1971).
3
" The process is
similar to the latitudinal differences within the range of a
given species.
Highly mobile species that depend upon temperature
synchrony among widely different regions or environments
for various phases of the reproductive or rearing cycle (e.g.,
anadromous salmonids or aquatic insects) could be faced
with dangers of dis-synchrony if one area is warmed, but
another is not. Poor long-term success of one year class of
Fraser River (British Columbia) sockeye salmon
(Oncorhyn-
chus nerka)
was attributed to early (and highly successful)
fry production and emigration during an abnormally warm
summer followed by unsuccessful, premature feeding
activity in the cold and still unproductive estuary (Vernon
1958).
3
" Anadromous species are able, in some cases, (see
studies of eulachon
(Thaleichthys pacificus)
by Smith and
Saalfeld 1955)
3
" to modify their migrations and spawning
to coincide with the proper temperatures whenever and
wherever they occur.
Rates of embryonic development that could lead to pre-
mature hatching are determined by temperatures of the
microhabitat of the embryo. Temperatures of the micro-
habitat may be quite different from those of the remainder
of the waterbody. For example, a thermal effluent at the
temperature of maximum water density (approximately
4 C) can sink in a lake whose surface water temperature
is colder (Hoglund and Spigarelli, 1972).290
Incubating
eggs of such species as lake trout
(Salvelinus namaycush)
and
various coregonids on the lake bottom may be intermittently
exposed to temperatures warmer than normal. Hatching
may be advanced to dates that are too early for survival of
the fry in their nursery areas. Hoglund and Spigarelli
1972,290 using temperature data from a sinking plume in
Lake Michigan, theorized that if lake herring
(Coregonus
artedii)
eggs had been incubated at the location of one of
their temperature sensors, the fry would have hatched
seven days early. Thermal limitations must, therefore, apply
at the proper location for the particular species or life stage
to be protected.
Recommendations
After their specific limiting temperatures and
exposure times have been determined by studies
tailored to local conditions, the reproductive ac-
tivity of selected species will be protected in areas
where:
•
periods required for gonad growth and gamete
maturation are preserved;
•
no temperature differentials are created that
block spawning migrations, although some delay
or advancement of timing based upon local con-
ditions may be tolerated;
41
Heat and Temperature/165
•
temperatures are not raised to a level at which
necessary spawning or incubation temperatures
of winter-spawning species cannot occur;
•
sharp temperature changes are not induced in
spawning areas, either in mixing zones or in
mixed water bodies (the thermal and geographic
limits to such changes will be dependent upon
local requirements of species, including the
spawning microhabitat, e.g., bottom gravels,
littoral zone, and surface strata);
•
timing of reproductive events is not altered to
the extent that synchrony is broken where repro-
duction or rearing of certain life stages is shown
to be dependent upon cyclic food sources or other
factors at remote locations.
•
normal patterns of gradual temperature changes
throughout the year are maintained.
These requirements should supersede all others
during times when they apply.
CHANGES IN STRUCTURE OF AQUATIC COMMUNITIES
Significant change in temperature or in thermal patterns
over a period of time may cause some change in the com-
position of aquatic communities (i.e., the species represented
and the numbers of individuals in each species). This has
been documented by field studies at power plants (Trembley
1956-1960) 321
and by laboratory investigations (McIntyre
1968). 3 °3
Allowing temperature changes to alter significantly
the community structure in natural waters may be detri-
mental, even though species of direct importance to man
are not eliminated.
The limits of allowable change in species diversity due to
temperature changes should not differ from those applicable
to any other pollutant. This general topic is treated in
detail. in reviews by others (Brookhaven National Lab.
1969) 258
and is discussed in Appendix Community
Structure and Diversity Indices, p. 408.
NUISANCE ORGANISMS
Alteration of aquatic communities by the addition of heat
may occasionally result in growths of nuisance organisms
provided that other environmental conditions essential to
such growths (e.g., nutrients) exist. Poltoracka (1968)3"
documented the growth stimulation of plankton in an
artificially heated small lake; Trembley (19659 re-
ported dense growths of attached algae in the discharge
canal and shallow discharge plume of a power station (where
the algae broke loose periodically releasing decomposing
organic matter to the receiving water). Other instances of
algal growths in effluent channels of power stations were
reviewed by Coutant (1970c).269
Changed thermal patterns (e.g., in stratified lakes) may
greatly alter the seasonal appearances of nuisance algal
growths even though the temperature changes are induced
by altered circulation patterns (e.g., artificial destratifica.
tion). Dense growths of plankton have been retarded ill
some instances and stimulated in others (Fast 1968 ;
275
and
unpublished data
1971).325
Data on temperature limits or thermal distributions in
which nuisance growths will be produced are not presently
available due in part to the complex interactions with other-
growth stimulants. There is not sufficient evidence to say
that any temperature increase will necessarily result in
increased nuisance organisms. Careful evaluation of local
conditions is required for any reasonable prediction of
effect.
Recommendation
Nuisance growths of organisms ,may develop
where there are increases in temperature or alter-
ations of the temporal or spatial distribution
of
heat
in water. There should be careful evaluation'
of all factors contributing to nuisance growths at
any site before establishment of thermal limits
based upon this response, and temperature limits
should be set in conjunction with restrictions on
;,4
other factors (see the discussion of Eutrophication:
and Nutrients in Section
I).
CONCLUSIONS
Recommendations for temperature limits to protect
aquatic life consist of the following two upper limits for any
time of the year (Figure 111-6).
1. One limit consists of a maximum weekly average
temperature that:
(a) in the warmer months (e.g., April through
October in the North, and March through
November in the South) is one third of the range
between the optimum temperature and the
ultimate upper incipient lethal temperature for the
most sensitive important species (or appropriate
life stage) that is normally found at that location at
that time; or
(b)
in the cooler months (e.g., mid-October to mid-
April in the North, and December to February in
the South) is that elevated temperature from which
important species die when that elevated tem-
perature is suddenly dropped to the normal
ambient temperature, with the limit being the
acclimation temperature (minus a 2 C safety
factor), when the lower incipient lethal tempera-
ture equals the normal ambient water temperature
(in some regions this limit may also be applicable
in summer) ; or
(c)
during reproduction seasons (generally April-June
and September-October in the North, and March-
May and October-November in the South) is that
42
I
Time-Temperature History
F
for Short Exposures
Time-Temperature
Limits for
Short Exposures
/ "`•
\_
,
/A
I
/
f- )
,
Seasons of
Reproduction
Requirements
c
-■A?
Maximum
(Based on
Weekly
species or
Average,
community)
Summer
Maximum
Weekly
Average,
Winter
32
86
Maximum Weekly
Average, Winter
166/Sectio
n
III—Freshwater Aquatic Life and Wildlife
temperature that meets specific site requirements
for successful migration, spawning, egg incubation,
fry rearing, and other reproductive functions of
important species; or
(d) at a specific site is found necessary to preserve
normal species diversity or prevent undesirable
growths of nuisance organisms.
2. The second limit is the time-dependent maximum
temperature for short exposures as given by the species-
specific equation:
1 >
time
10fa±b(temp.+2)]
Local requirements for reproduction should supersede
all other requirements when they are applicable. Detailed
ecological analysis of both natural and than-modified
aquatic environments is necessary to ascertain when these
requirements should apply.
USE OF TEMPERATURE CRITERIA
A hypothetical electric power station using lake water for
cooling is illustrated as a typical example in Figure 111-7.
This discussion concerns the application of thermal criteria
to this typical situation.
The size of the power station is 1,000 megawatts electric
(MW
e
) if nuclear, or 1,700 MW
e
if fossil-fueled (oil, coal,
gas) ; and it releases 6.8 billion British Thermal Units
(BTU) per hour to the aquatic environment. This size is
representative of power stations currently being installed.
Temperature rise at the condensers would be 20 F with
cooling water flowing at the rate of 1,520 cubic feet/second
(ft
3
/sec) or 682,000 gallons/minute. Flow could be in-
creased to reduce temperature rise.
The schematic Figure 111-7 is drawn with two alternative
discharge arrangements to illustrate the extent to which
design features affect thermal impacts upon aquatic life
20
EL'
10
0
C
30
J?
F?
M?
A
M
?
J
?
J
?
S?
0
N?
D
Annual Calendar
FIGURE III-6—Schematic Summary
of Thermal
Criteria
Power
Plant
Rapid Dilution
D
80
70
60
E
50
40
30
20
J F M A M
J
Months
A SON
D
Plume Scale
0?
5000
I
i i
1
Feet
Modified
— Temperature
(Example)
Heat and Temperature/167
FIGURE III-7—Hypothetical Power Plant Site For Application of Water Temperature Criteria
168/Section HI—Freshwater Aquatic Life and Wildlife
Warm condenser water can be carried from the station to
the lake by (a) a pipe carrying water at a high flow velocity
or (b) a canal in which the warm water flows slowly. There
is little cooling in a canal, as measurements at several
existing power stations have shown. Water can be released
to the lake by using any of several combinations of water
velocity and volume (i.e., number of outlets) or outlet
dimensions and locations. These design features largely
determine the configuration of the thermal plumes illus-
trated in Figure 111-7 resulting from either rapid dilution
with lake water or from slow release as a surface layer. The
isotherms were placed according to computer simulation
of thermal discharges (Pritchard 1971)
312
and represent a
condition without lake currents to aid mixing.
Exact configuration of an actual plume depends upon
many factors (some of which change seasonally or even
hourly) such as local patterns of currents, wind, and bottom
and shore topography.. •
Analytical Steps
Perspective of the organisms in the water body and of the
pertinent non-biological considerations (chemical, hy-
drological, hydraulic) is an essential beginning. This
perspective requires a certain amount of literature survey
or on site study if the information is not well known. Two
steps are particularly important:
1. identification of, the important species and com-
munity (primary production, species diversity, etc.) that are
relevant to this site; and
2.
determination of life patterns of the important species
(seasonal distribution, migrations, spawning areas, nursery
and rearing areas, sites of commercial or sport fisheries).
This information should include as much specific informa-
tion on thermal requirements as it is possible to obtain
from the literature.
Other steps relate the life patterns and environmental
requirements of the biota to the sources of potential thermal
damage from the power plant. These steps can be identified
with specific areas in Figure 111-7.
Aquatic Areas Sensitive to Temperature Change
Five principal areas offer potential for biological damage
from thermal changes, labeled A-E on Figure 111-7. (There
are other areas associated with mechanical or chemical
effects that cannot be treated here; see the index.)
Area A The cooling water as it passes through the intake,
intake piping (A 1
), condensers, discharge piping
(A2
) or canal (A'
2
), and thermal plume (A
3 or
A'3
), carrying with it small organisms (such as
phytoplankton, zooplankton, invertebrate larvae,
and fish eggs or larvae). Organisms receive a
thermal shock to the full 20 F above ambient
temperature with a duration that depends upon
the rate of water flow and the temperature drop
in the plume.
Area B Water of the plume alone that entrains both
small and larger organisms (including small fish)
as it is diluted (B or B'). Organisms receive
thermal shocks from temperatures ranging from
the discharge to the ambient temperature, de-
pending upon where they are entrained.
Area C Benthic environment where bottom organisms
(including fish eggs) can be heated chronically or
periodically by the thermal plume (C or C').
Area D The slightly warmed mixed water body (or large
segment of it) where all organisms experience a
slightly warmer average temperature (D).
Area E The discharge canal in which resident or seasonal
populations reside at abnormally high tempera-
tures (E).
Cooling Water Entrainment
It is not adequate to consider only thermal criteria for
water bodies alone when large numbers of aquatic organisms
may be pumped through a power plant. The probability
of an organism being pumped through will depend upon
the ratio of the volume of cooling water in the plant to the
volume in the lake (or to the volume passing the plant in a
river or tidal fresh water). Tidal environments (both
freshwater and saline) offer greater potential for entrain-
ment than is apparent, since the same water mass will
move back and forth past the plant many times during the
lifetime of pelagic residence time of most organisms.
Thermal shocks that could be experienced by organisms
entrained at .the hypothetical power station are shown in
Figure III-8.
Detrimental effects of thermal exposures received during
entrainment can be judged by using the following equation
for short-term exposures to extreme temperatures :
General criterion:
1
—
>
10(a±b(temp.+2)1
Values for a and b in the equation for the species of aquatic
organisms that are likely to be pumped with cooling water
may be obtained from Appendix II, or the data may be
obtained using the methods of Brett (1952).
25' The prevailing
intake temperature would determine the acclimation
temperature to be selected from the table.
For example, juvenile largemouth bass may frequent the
near-shore waters of this lake and be drawn into the intake.
To determine whether the hypothetical thermal discharges
(Figure 111-7) would be detrimental for juvenile bass, the
following analysis can be made (assuming, for example,
that the lake is in Wisconsin where these basic data for bass
are available):
Criterion for juvenile bass (Wisconsin) when intake
time
12
12
Canal
Condenser
Piping
10
Canal Plume
Intake Piping
2
Dilution Plume
Intake
0
4
8
1
1
1
12
16
20
24
28
32
1
36
0
40
1>
— 1
0[34.3649-0.97
89(temp.+2)]
time
Heat and Temperature/169
20
Time After Initial Heating (hrs.)
(C°
Modified after Coutant 1970c269
FIGURE III-8—Time Course of Temperature Change
in
Cooling Water Passing Through the Example Power Station with
Two Alternate Discharges. The
Canal
Is Assumed to Flow at a Rate of 3 Ft. Per Sec.
temperature (acclimation) is 70 F (21.11 C). (Data
from Appendix II-C).
Canal
Criterion applied to entrainment to end of discharge
canal (discharge temperature is 70 F plus the 20 degree
rise in the condensers or 90 F (32.22 C). The thermal
plume would provide additional exposure above the
lethal threshold, minus 2 C (29.5 C or 85.1 F) of more
than four hours.
1>
— 10[34.3649-0.9789(32.22+2)]
1 >8.15
Conclusion:
Juvenile bass would not survive to the end of the
discharge canal.
Dilution
Criterion applied to entrainment in the system em-
ploying rapid dilution.
1.2
1 —
10(34.3649-0.9789(32.22+2.0)]
1.2
1>
7.36
Travel time in piping to discharge is assumed to be
1 min., and temperature drop to below the lethal
threshold minus 2 C (29.5 C or 85.1 F) is about 10 sec.
(Pritchard, 1970.312
Conclusion
Juvenile bass would survive this thermal exposure:
1>0.1630
By using the equation in the following form,
log (time)=a—{-b (temp.+2)
the length of time that bass could barely survive the
expected temperature rise could be calculated, thus
allowing selection of an appropriate discharge system.
For example:
log (time) =34.3649-0.9789 (34.22)
log (time) =0.8669
time =7.36
60
G
1 701 Section III—Freshwater Aquatic Life and Wildlife
This would be about 1,325 feet of canal flowing at
3 ft/sec.
It is apparent that a long discharge canal, a nonrecircu-
lating cooling pond, a very long offshore Moe, or delayed
dilution m a mixing zone (such as the one promoting surface
cooling) could prolong the duration of exposure of pumped
organism
s
and thereby increase the likelihood of damage to
them. Precise information on the travel times of the cooling
water in the discharge system is needed to conduct this
analysis.
The calculations have ignored changing temperatures in
the thermal plume, because the canal alone was lethal, and
cooling in the plume with rapid dilution was so rapid that
the additional exposure was only for 10 seconds (assumed to
be at the discharge temperature the whole time). There
may be other circumstances under which the effect of
decrealing exposure temperature in the plume may be
of interest.
Effects of changing temperatures in the plume can be
estimated by summing the effects of incremental exposures
for short time periods (Fry et al. 1946
281
). For example, the
surface cooling plume of Figures 111-7 and 111-8 could be
considered to be composed of several short time spans, each
with an average temperature, until the temperature had
dropped to the upper lethal threshold minus 2 C for the
juvenile bass. Each time period would be calculated as if
it were a single exposure, and the calculated values for all
time periods would be summed and compared with unity,
as follows :
timer
time2
time„
10 [a-l-b(temp./.+2)1
10
[a+b(temn . 2
+2)]
10
Ca+b(temp.”1-2)]
The surface cooling plume of Figure 111-6 (exclusive of
the canal) could be considered to consist of 15 min it
89.7 F (32.06 C), 15 min at 89.2 F (31.78 C), 15 min at
88.7 F (31.4 C), 15 min at 88.2 F (31.22 C), 15 min at
87.8 F (31.00 C), until the lethal threshold for 70 F acclima-
tion minus 2 C (85.1 F) was reached. The calculation would
proceed as follows:
1
1 >
15
—
10134.3049-0. 97 89(32. 06+2)]
15
+
10134.3649-0.8 789(31.7 8+2)1 +
• • •
In this case, the bass would not survive through the first
15
-minute period. In other such calculations, several steps
would have to be summed before unity was reached (if not
r
eached, the plume would not be detrimental).
En
t
r
ainment in the Plume
Organisms mixed with the thermal plume during dilution
will also receive thermal shocks, although the maximum
tem
peratures will generally be less than the discharge
temperature. The number of organisms affected to some
degree may be significantly greater than the numbers
actually pumped through the plant. The route of maximum
thermal exposure for each plume is indicated in Figure
111-7
by a dashed line. This route should be analyzed to
determine the maximum reproducible effect.
Detrimental effects of these exposures can also be judged
by using the criterion for short-term exposures to extreme
temperatures. The analytical steps were outlined above for
estimating the effects on organisms that pass through the
thermal plume portions of the entrainment thermal pattern.
There would have been no mortalities of the largemouth
bass from entrainment in the plume with rapid dilution, due
to the short duration of exposure (about 10 seconds). Any
bass that were entrained in the near-shore portions of the
larger plume,
and
remained in it, would have died in less
than 15 minutes.
Bottom Organisms Impacted
by
the Plume
Bottom communities of invertebrates, algae, rooted
aquatic plants, and many incubating fish eggs can be
exposed to warm plume water, particularly in shallow
environments. In some circumstances the warming can be
continuous, in others it can be intermittent due to changes
in plume configuration with changes in currents, winds, or
other factors. Clearly a thermal plume that stratifies and
occupies only the upper part of the water column will have
least effect on bottom biota.
Several approaches are useful in evaluating effects on the
community. Some have predictive capability, while others
are suitable largely for identifying effects after they have
occurred. The criterion for short-term exposures identified
relatively brief periods of detrimental high temperatures.
Instead of the organism passing through zones of elevated
temperatures, as in the previous examples, the organism is
sedentary, and the thermal pulse passes over it. Developing
fish eggs may be very sensitive to such changes. A brief
pulse of high temperature that kills large numbers of orga-
nisms may affect a bottom area for time periods far longer
than the immediate exposure time. Repeated sublethal ex-
posures may also be detrimental, although the process is
more complex than straight-forward summation. Analysis
of single exposures proceeds exactly as described for plume
entrainment.
The criterion for prolonged exposures is more generally
applicable. The maximum tolerable weekly average tem-
perature may be determined by the organisms present and
the phase of their life cycle. In May, for example, the
maximum heat tolerance temperature for the community
may be determined by incubating fish eggs or fish fry on the
bottom. In July it may be determined by the important
resident invertebrate species. A well-designed thermal dis-
charge should not require an extensive mixing zone where
these criteria are exempted. Special criteria for reproductive
processes may have to be applied, although thermal dis-
Heat and Temperature/171
charges should be located so that zones important for
reproduction—migration, spawning, incubation—are not
used.
Criteria for species diversity provide a useful tool for
identifying effects of thermal changes after they have
occurred, particularly the effects of subtle changes that are
a result of community interactions rather than physiological
responses by one or more major species. Further research
may identify critical temperatures or sequences of tem-
perature changes that cannot be exceeded and may thereby
provide a predictive capability as well. (See Appendix
II–B.)
Mixed Water Body (or major region thereof)
This is the region most commonly considered in es-
tablishing water quality standards, for it generally includes
the major area of the water body. Here the results of thermal
additions are observed as small temperature increases over a
large area (instead of high temperatures locally at the dis-
charge point), and all heat sources become integrated into
the normal annual temperature cycle (Figure 111-6 and
Figure 111-7 insert).
Detrimental high temperatures in this area (or parts of
it) are defined by the criteria for maximum temperatures
for prolonged exposure (warm and cool months) for the
most sensitive species or life stage occurring there, at each
time of year, and by the criteria for reproduction.
For example, in the lake with the hypothetical power
station, there may be 40 principal fish species, of which half
are considered important. These species have spawning
temperatures ranging from 5 to 6 C for the sauger
(Stizo-
stedion canadense)
to 26.7 C for the spotted bullhead
(Ictalurus
serracanthus).
They also have a similar range of temperatures
' required for egg incubation, and a range of maximum
temperatures for prolonged exposures of juveniles and
adults. The requirements, however, may be met any time
within normal time spans, such as January 1 to 24 for sauger
spawning, and March 25 to April 29 for smallmouth bass
spawning. Maximum temperatures for prolonged exposures
may increase steadily throughout a spring period. To
predict effects of thermal discharges the pertinent tempera-
tures for reproductive activities and maximum temperatures
for each life stage can be plotted over a 12-month perio
such as shown in Fig. 111-6. A maximum annual tempera-
ture curve can become apparent when sufficient biological
data are available. Mount (1970)
305
gives an example of
this type of analysis.
Discharge Canal
Canals or embayments that carry nearly undiluted
condenser cooling water can develop biological communities
that are atypical of normal seasonal communities. Interest
in these areas does not generally derive from concern for a
balanced ecosystem, but rather from effects that the altered
communities can have on the entire aquatic ecosystem.
The general criteria for nuisance organisms may be
4
applicable. In• the discharge canals of some existing power
stations, extensive mats of temperature-tolerant blue-green
algae grow and periodically break away,. adding a decom-
posing organic matter to 'the nearby shorelines.
The winter criterion for maximum temperatures for
prolonged exposures identifies the potential for fish kills due
to rapid decreases in temperature. During cold seasons
particularly, fish are attracted to warmer water of an
enclosed area, such as a discharge canal. Large numbers
may reside there for sufficiently long periods to become
metabolically acclimated to the warm water. For any
acclimation temperature there is a minimum temperature
to which the species can be cooled rapidly and still survive
(lower incipient lethal temperature). These numerical
combinations, where data are available, are found in
Appendix
II
–C. There would be 50 per cent mortality, for
example, if largemouth bass acclimated in a discharge
canal to 20 C, were cooled to 5.5 C or below. If normal
winter ambient temperature is less than 5.5 C, then the
winter maximum should be below 20 C, perhaps nearer
15 C. If it is difficult to maintain the lower temperatures,
fish should be excluded from the area.
HEAT AND TEMPERATURE
242
Allen, K. 0. and K. Strawn (1968), Heat tolerance of channel
catfish
Ictalurus punctatus,
in
Proceedings of the 21st annual con-
ference of the Southeastern Association of Game and Fish Commissioners
(The Association, Columbia, South Carolina), pp. 399-411.
241
Anderson, R. 0. (195
.
9), The influence of season and temperature
on the growth of the bluegill
(Lepomis macrochirus).
Ph.D. thesis,
University of Michigan, Horace H. Rackham School of Graduate
Studies. 133 p.
242
Andrews,
J.
W. and R. R. Stickney (1972), Interaction of feeding
rates and environmental temperature of growth, food conversion,
and body composition of channel catfish.
Trans. Amer. Fish. Soc.
101(0:94-99.
242
Ansell, A. D., 1968. The Rate of Growth of the hard clam
Mer-
cenaria mercenaria
(L) throughout the geographical range. Conseil
permanent international pour l'exploration de la mer. 31:(3)
364-409.
244
Baldwin, N. S. (1957), Food consumption and growth of brook
trout at different temperatures.
Trans. Amer. Fish. Soc.
86:323-
246
C. D., C. C. Coutant, and E. F. Prentice (1971),
Experi-
mental drifts of juvenile salmonids through effluent discharges at Hanford.
Part II. 1969 drifts and conclusions
[USAEC BNWL-1527] (Bat-
telle-Northwest, Richland, Washington), 61 p.
246
Beyerle, G. B. and
Cooper, E. L. (1960), Growth of brown trout
in selected Pennsylvania streams,
Trans. American Fisheries Society
89(3): 255-262.
247
Bishai, H. M. (1960), Upper lethal temperatures for larval sal-
monids.
j.
Cons. Perma. Int. Explor. Mer
25(2):129-133. .
242
Black, E. C. (1953), Upper lethal temperatures of some British
Columbia freshwater fishes.
J. Fish. Res. Bd. Canada
10(4):196-
210.
249
C. I. (1937), Calculation of the time-mortality curve.
Ann.
Appl. Biol.
24:815-852.
262
Breder, C. M. and D. E. Rosen (1966),
Modes of reproduction in
fishes
(The Natural History Press, New York), 941 p.
261
J. R. (1941), Tempering versus acclimation in the planting
of speckled trout.
Trans. Amer. Fish. Soc.
70:397-403.
262
Brett, J. R. (1952), Temperature tolerance in young Pacific sal-
mon, genus
Oncorhynchus. J. Fish. Res. Bd. Canada
9:265-323.
2 g
2
?
T A /InCeN
Literature Cited/205
254
Brett, J. R. (1960), Thermal requirements of fish—three decades
of study, in
Biological problems of water pollution,
C. M. Tarzwell,
ed. (U.S. Department of Health, Education and Welfare, Robert
A. Taft Sanitary Engineering Center, Cincinnati, Ohio), pp.
110-117.
2 "
Brett, J. R. (1970), Temperature-animals-fishes, in
Marine
ecology,
0. Kinne, ed. (John Wiley & Sons, New York), vol. I,
pp. 515-560.
266 Brett, J. R. (1971), Energetic responses of salmon to temperature.
A study of some thermal relations in the physiology and fresh
water ecology of sockeye salmon
(Oncorhynchus nerka). Amer. Zool.
11(1):99-113.
252
Brett, J. R., J. E. Shelbourn, and C. T. Shoop (1969), Growth
rate and body composition of fingerling sockeye salmon,
Oncor-
hynchus nerka,
in relation to temperature and ration size.
5.
Fish.
Res. Bd. Canada
26(9):2363-2394.
258
Brookhaven National Laboratory (1969), Diversity and stability
in ecological systems.
Brookhaven Symposia in Biology
22:264 pp.
259
Bullock,.T.
H. (1955), Compensation for temperature-in the metab-
olism and activity of poikilotherms.
Biol. Rev.
(Cambridge) 30(3):
311-342.
269
Burdick, G. E., H. J. Dean, E. J. Harris, J. Skea, C. Frisa and C.
Sweeney (1968), Methoxychlor as a blackfly larvicide: persistence
of its residues in fish and its effect on stream arthropods.
N.Y.
Fish. Game].
15(2):121-142.
261
Cairns, J., Jr. (1968), We're in hot water.
Scientist and Citizen
10(8)
187-198.
262
Churchill, M. A: and T. A. Wojtalik (1969), Effects of heated dis-
charges on the aquatic environment: the TVA experience, in
Prooceedings American Power Conference
(Tennessee Valley Authority,
Chattanooga), vol. 31, pp. 360-368.
263 Clark, J.:
R. (1969), Thermal pollution and aquatic life.
Sci. Amer.
220(3).:18-27.
264
Cooper, E. L. (1953), Periodicity of growth and change of condi-
tion of brook trout
(Salvelinus fontinalis)
in three Michigan trout
streams. Copeia 1953(2):107-114.
266
Coutant, C. C. (1968), Thermal pollution—biological effects: a
review of the literature of 1967.5.
Water Pollut. Contr. Fed.
40(6):
1047-1052.
"6
Coutant, C. C. (1969), Thermal pollution-biological effects: a
review of the literature of 1968.
Water Pollut. Contr. Fed.
41(6):
1036-1053.
267
Coutant, C. C. (1970a), Thermal pollution—biological effects: a
review of the literature of 1969.5.
Water Pollut. Contr. Fed.
42(6):
1025-1057. •
266
C. C. (1970b),
Thermal resistance of adult coho
(Oncor-
hynchus kisutch)
and jack chinook (0.
tshawytscha)
salmon, and the
adult steelhead trout
(Salmo gairdnerii)
from the Columbia River
[SEC BNWL-1508 ] Battelle-Northwest, Richland, Washington,
24 p.
269 Coutant, C. C. (1970c), Biological aspects of thermal pollution. I.
Entrainment and discharge canal effects.
CRC Critical Rev. En-
viron. Contr.
1 (3):341-381.
27
°
Coutant, C. C. (1971), Thermal pollution-biological effects.
3.
Water Pollut. Contr. Fed.
43(6):1292-1334.
221
Coutant, C. C. and R. M. Steele (1968), Effect of temperature on
the development rate of bottom organisms, in Pacific Northwest
Laboratory Annual Report for 1967' to USAEC Division of
Biology and Medicine, vol. 1, Biological Sciences, Thompson,
R. C., P. Teal and E. G. Swezes, eds. [BNWL-714] Battelle-
Northwest, Richland, Washington.
222D
oudoroff,
P. and D. L. Shumway (1970),
Dissolved oxygen require-
ments of freshwater fishes
[Food and Agricultural Organization
fisheries technical paper 86] (FAO, Rome), 291 p.
3
Ebel, W. J., E. M. Dawley, and B. Monk (1970), Thermal tolerance
of juvenile Pacific salmon in relation to supersaturation of nitro-
gen gas.
Fish. Bull.
69 (4):833-843.
206/ Section III—Freshwater Aquatic Life and Wildlife
274
Edsall, T. A. and P. J. Colby (1970), Temperature tolerance of
young-of-the-year Cisco,
Coregonus artedii. Transactions of American
Fisheries Society
99:(3)526-531.
"'Fast, A. W. (1968), Artificial destratification of El Capitan reser-
voir by aeration. I. Effects on chemical and physical parameters.
Calif. Dep. Fish Game Fish Bull.
no. 141,97 p.
276
Fry, F. E. J. (1947), Effects of the environment on animal activity.
Univ. of Toronto Stud. Biol. Ser. No.
55 Publ.
Ont. Fish. Resh. Lab.
No.
68:1-62.
272 Fry, F. E. J. (1951), Some environmental relations of the speckled
trout
(Salvelinas fontinalis).
Proc. Northeast. Atlantic Fisheries
Conf. May, 1951.
"
8
Fry, F. E. J. (1964), Animals in aquatic environments: fishes
temperature effects (Chapter 44) Handbook of Physiology,
Section 4: Adaptation to the Environment.
Amer. Physiol. Soc.,
Washington, D. C.
"8
Fry, F. E. J. (1967), Responses of vertebrate poikilotherms to
temperature [review ], in
Thermobiology,
A. H. Rose, ed. (Aca-
demic Press, New York), pp. 375-409.
280 Fry,•F. E. J., J. R. Bi-ett, and G. H. Clawson (1942), Lethal limits
of temperature for young goldfish.
Rev. Can. Biol.
1(1):50-56.
981 Fry, F. E.
5.,
J. S. Hart, and K. F. Walker (1946), Lethal tempera-
ture relations for a sample of young speckled trout,
Savelinus
fontinalis
[University of Toronto biology series no. 54] (The
University of Toronto Press, Toronto), pp. 9-35.
2 " Gammon, J. R. (1970),
Aquatic life survey of the Wabash River, with
special reference to the effects of thermal effluents on populations of micro-
invertebrates and fi
sh, 7967-1969
(DePauw University, Zoology De-
partment, Greencastle, Indiana), 65 p.
282
Gibson, E. S. and F. E. J. Fry (1954), The performance of the lake
trout,
Salvelirws namaycush,
at various levels of temperature and
oxygen pressure. Can.
3
•ool.
32(3):252-260.
284 Graham, J. M. (1949), Some effects of temperature and oxygen
pressure on the metabolism and activity of the speckled trout
Salvelinus fonlinalis. Can. 3. Res
(D) 27:270-288.
26s
?J. S. (1947), Lethal temperature relations of certain fish in
the Toronto region.
Trans. Roy. Soc. Can. (Sec.
5) 41:57-71.
286 Hart, J. S. (1952),
Geographical variations of some physiological and
morphological characters in certain freshwater fish.
(University of
Toronto biology series no. 60) (The University of Toronto Press,
Toronto), 79 p.
287
A. L. (1961), A review of the nature and extent of damage
caused by oil pollution at sea.
Trans. N. Am. Wild!. and Nat. Re-
sources Conf.
26:343-355.
288 Heinle, D. R. (1969), Temperature and zooplankton.
Chesapeake
Sci.
10(3-4):186-209.
289
Hoff, J. G. and J. R. Westman (1966), The temperature tolerances
of three species of marine fishes.
3. Mar. Res.
24(2):131-140.
280 Hoglund, B. and S. A. Spigarelli (1972), Studies of the sinking
plume phenomenon. Argonne National Lab., Center for Envir.
Stud., Argonne, Ill.
291
W. B. II and R. E. Pearson (1972), Growth, tempera-
ture requirements and lower lethal temperature for juvenile
smallmouth bass
(Micropterus dolomieu
Lacepede). Draft manu-
script, U.S. National Water Quality Laboratory, Duluth, Minn.
282
R. T. and M. 0. Synoground (1970),
Effect of Hanford Plant
operations on the temperature of the Columbia River 1964 to the present
[BNWL-1345 ] (Battelle-Northwest, Richland, Washington),
various paging.
"3
Jensen, A. L. (1971), The effect of increased mortality on the
young in a population of brook trout: a theoretical analysis.
Trans. Amer. Fish. Soc.
100(3)1456-459.
294
Kennedy, V. S. and J. A. Mihursky (1967),
Bibliography on the ef-
fects of temperature in the aquatic environment
[Contribution 326]
(University of Maryland, Natural Resources Institute, College
Park) 89 p.
"5
Kinne, 0. (1963), The effects of temperature and salinity on
marine and brackish water animals. I. temperature.
Oceanogr.
Mar. Biol. Annul Rev.
1:301-340.
296 Kinne, 0. (1970), Temperature—animals—invertebrates, in
Marine ecology, 0.
Kinne, ed. ( John Wiley & Sons, New York),
vol. 1, pp. 407-514.
297 Kramer, R. H. and L. L. Smith Jr. (1960). First year growth of
the largemouth bass,
Micropterns salrnoides
(Laccpdc) and some
related ecological factors.
Transactions American Fisheries Society
89(2):222-233.
298 Krenkel, P. A. and F. L. Parker, eds. (1969),
Biological aspects of
thermal pollution
(Vanderbilt University Press, Nashville, Ten-.
nessee), 407 p.
299 Lawler, G. H. (1965), Fluctuations in the success of year-classes of
white-fish populations with special reference to Lake Erie.
3.
Fish. Res. Bd. Canada
22(5):1197-1227.
300 Lemke, A. L. (1970), Lethal effects of various rates of temperature
increase on
Gammarus pseudolimnaeus
and
Hydropsyche bctteni
with
notes on other species. U.S. National Water Quality Laboratory;
Duluth, Minnesota.
301
McCornish, T. S. (1971), Laboratory experiments on growth and
food conversion by the bluegill. Ph.D. dissertation, Univ. of
Missouri, Columbia, Mo.
•
302
McCormick, J. H. et, al. (1971), Temperature requirements for
growth and survival for Larvae Ciscos
(Coregonus artedii). lour.
Fish. Res. Bd. Canada
28:924.
303
McIntire, C. D. (1968), Physiological-ecological studies of benthic
algae in laboratory streams.
5.
Water Pollul. Contr. Fed.
40(11
part 1):1940-1952.
304
Merriman, D., et al. (1965), The Connecticut River investigation,
1965-1972. (A series of semi-annual progress reports). Connecti-
cut Yankee Atomic Power Company, Haddar, Connecticut.
395 Mount, D. I. (1970), Statement before hearing before the Joint
Committee on Atomic Energy, Congress of the United States,
Ninety-First Congress, first session [on environmental effects of
producing electric power.] part 1, pp. 356-373.
398
Narver, D. W. (1970), Diel vertical movements and feeding of
underyearling sockeye salmon and the limnetic zooplankton in
Babine Lake, British Columbia.
5. Fish. Res. Bd. Canada
27(2):
281-316.
397
Nebeker, A. V. (1971), Effect of temperature at different altitudes
on the emergence of aquatic insects from a single stream.
J.
Kans. Entomol. Soc.
44(1):26-35.
3
"
Parker, F
P I.. and P. A. Krenkel, eds. (1969),
Engineering aspects
of thermal pollution
(Vanderbilt University Press, Nashville, Ten-
nessee), 351 p.
399
Peek, F. W. (1965). Growth studies of laboratory and wild popu-
lation samples of srnallmouth bass
(Micropterus dolomieu
Lacepede)
with applications to mass marking of fishes. M.S. Thesis, Univ.
of Arkansas, Fayetteville.
329
Pennsylvania Fish Commission (1971), Water pollution report no.
4170.
311
Poltoracka, J. (1968), [Specific composition of phytoplankton in a
lake warmed by waste water from a thermoelectric plant and
lakes with normal temperature.]
Acta. Soc. Bot. Pol.
37(2):297-
325.
312
Pritchard, D. W. (1971), Design and siting criteria for once-
through cooling systems. Presented at the American Institute of
Chemical Engineers 68th annual meeting, 2 March 1971,
Houston, Texas.
313
Raney, E. C. and B. W. Menzel (1969),
Heated e
f
fluents and effects
on aquatic life with emphasis on fishes: a bibliography,
38th ed. (U.S.
Department of the Interior, Water Resources Information Center,
Washington, D.C.), 469 p.
314
Robinson, J. G. (1968),
Fish mortality report, Lake Michigan, Port
Sheldon, August 29, 1968
(Michigan Water Resources Commis-
sion, Lansing); 2 p.
318
Robinson, J. G. (1970), Fish mortality report, Lake Michigan,
Port Sheldon. Michigan Water Resources Commission, Lansing,
Michigan.
318
Robinson, J. G. (1970), Fish mortality report, Lake Michigan,
Port Sheldon. Michigan Water Resources Commission Lansing,
Michigan.
37
Smith, W. E. and R. W. Saalfeld (1955), Studies on Columbia
River smelt
Thaleichthys pacificus
(Richardson).
Wash. Dep. Fish.
Fish. Res. Pap.
1(3) :1-24.
2"
Smith, S. H. (1964), Status of the deepwater cisco population of
Lake Michigan.
Trans. Amer. Fish. Soc.
93(2):155-163.
3"
Strawn, K. (1961), Growth of largemouth bass fry at various
temperatures.
Trans. Amer. Fish. Soc.
90:334-335.
3
20
Strawn, K. (1970), Beneficial uses of warm water discharges in
surface waters.
In:
Electric power and thermal discharges:
thermal considerations in the production of electric power, M.
Eisenbud and G. Gleason (eds.) pp. 143-156..
3"
Trembley, F. J. (1965), Effects of cooling water from steam-eke-
tric power plants on stream biota, in
Biological problems in 'paler
pollution. Third seminar,
C. M. Tarzwell, ed. (U.S. Department of
Health, Education and Welfare, Public Health Service, Division
of Water Supply and Pollution Control, Cincinnati, Ohio), pp.
334-345.
323
Vernon, E. H. (1958), An examination of factors affecting the
abundance of pink salmon in the Fraser River [Progress report
no. 5] (International Pacific Salmon Fisheries Commission, New
Westminster, British Columbia).
323
Wiebe, J. P. (1968), The effects of temperature and day length
on the reproductive physiology of the viviparous scaperch,
Cymalogaster aggregala
Gibbons.
Can. 5. Zool.
.46(6):1207-1219.
References Cited
324 Coutant, C. C.,
unpublished data,
(1971) Oak Ridge Laboratory,
Oak Ridge, Tennessee.
328
Fast, A. W. (1971), Effects of artificial aeration on lake ecology.
Ph.D. dissertation, Michigan State Univ., E. Lansing.
326 Fry, F. E. J.,
personal observation,
(1971) University of Toronto,
Ontario, Canada, Dept. of Zoology.
3 "
Jones, B.,
unpublished data,
(1971) National Water Quality Labora-
tory, Duluth, Minnesota.
328 National Water Quality Laboratory (1971)
unpublished data,
Duluth,
Minnesota.
329
Wojtalik, T. A.,
unpublished data,
(1971) Tennessee Valley Authority.
TI
or;
exi
AEU
bl
m
Adif
(d
(h
AIM
Brn
m
m
Cab
•
at
50
APPENDIX B*
THERMAL TABLES
THERMAL TABLES-Time-temperature relationships and lethal threshold temperatures for resistance of aquatic
organisms (principally fish) to extreme temperatures (from Coutant,
in press'
s
1972). Column headings, where
not
self-
explanato
ry
, are identified
in
footnotes. LD50 data obtained for single times only were included
only
when they amplified
temperature-time information.
Accfimation
?
log lime= a+b (temp.)
?
Data hmits?
Lethal
Stage/age?
Length
?
Weight?
. Sex?
Location?
Reference
?
Extreme ?
?
(°C)?
L050?
thresholdd
Temp .
?Time?b?
N
b?
r,?
(°C)
upper lower
Abodeldel sass- Adult
??
?
Northern Gun Heath,
W. G.?
Upper..
?
32?42.1705 -0.0914
?
3?
-0.9945 31.0 36.0
?
fi
g
s (Sargent
?
of California?
(1967)"
rnel0r)
Adinia mica
?
Adult.,??
?
Jefferson Co., Than and
?
Upper..?
35
?
(0 n
/eo)
.
21.9331 -0.4866
?
6?
-0.9930 43.0 40.5 ?
(diamond Killi-
?
Texas
?
Dunn?
35?
(5 Vol)' 27.7919 -0.6159
?
6?
-0.9841 43.5 41.0 ?
Ish)?
(1967)"?
35?
(10
0
yoe). 26.8121 -0.5899?
6?
-0.9329 43.5 41.0
?
35?
(20 0/0).
28.3930 -0.6290?
6
?
-0.9734
.
43.5 41.0 ?
•
Allarinops Ards Juvenile__ 6.0-6.2 cm.?
?
Wove, Calif. Doudoroll?
Upper..
?
18.0?
?
30.5(24)
(toltsmeit)
?
(194579
?
20?
42.2531 -1.2215
?
9
?
-0.9836 33.5
3r.5 ??
31.0
Lower..?
14.5?
7.6(24)
18.0
?
8.8(24)
20?
?
-0.4661?
0.3926
?
1
?
0.9765 11.0
?
5.0 ??
10.5
25.5?
?
13.5(24)
Swords titan- Larval?
17-34 mm?
?
Mined
?
Beaufort Har- Lewis (1965)"
?
Lower?
7.0
?
0.9611/ 0.2564
?
9?
0.5601 4.0 ??
5.0
..
‘
nes
(Atlantic
?
bor, North?
"?
10.0?0.7572?
0.2526 12?
0.9452?
5.0 -1.0
??
6.0
menhaden)
?
Carolina
?
12.5?
0.6602?
0.2786 12?
0.9852 5.5
?
?
>7.0
(36°N)?
II
?
15.0?
0.5615
?
0.2321
?
14?
0.9306?
1.0 ?
?
>8.0
" 20.0?
0.2620?
0.1817?
3?
0.9612?
4.0
?
Irmoortia
tyran-
Young-of-the- ?
?
?
Beaufort;
?
Lewis and Net-?
Upper?
21?
(5 Von) 57.9980 -0.1643
?
2?
?
35.0 34.0 ?
nut (Atlantic
?
year
?
N.C.?
tier (1968)"?27
?
(5 Von) 85.1837 -2,3521
?
2?
?
35.0 34.5
?
menhaden)
?
Lower?
16?
(26-30 0
/00)?
7.0?
3.0 ??
6.5
18?
(10 0
/0o) ........
?
7.0?
3.0 ??
6.5
IFF100111 4
hymn-
Yearling?
?
Beaufort,?
Lewis and Het-?
Upper
?
21?
(5
0
/00)
35.7158 -1.0458
?
3?
-0.9174 34
?
33
MIS
(Atlantic
?
N.C.?
tier (1968)
n
?22-23?
(4-6 11 /W
21.8013 -0.6342 10
?
-0.9216 35
?
31?
32.5
menhaden)
Crudes stratus Juvenile
?
?
2g a
ye?
Mined?
Commercial?
Fry, Brett, &?
Upper?
1-2?
?
21 (14) ?
(goldfish)
?
dealer?
Clawson?
10?
?
31 (14) ?
(Toronto)?
(1942)81
(and?
17?
?
34 (14)
Fry, Hart, &
?
24?
........
?
....
?
?
36 (14)
??
Walker,
?
32?
210213 -0.4523
?
2
?
?
41.0 39.0 39.2(14)
?
1946)13
?38?
21.9234 -0.4773
?
2?
?
43.0 41.0 41.0(14)
?
41.0
Lower?
19?
1.0(14)
?
24
?
?
5.0(14)
38
?........?
....???
15.5(14)??
Spada
Ca
tostamus corn. Adult (1-2 yr)
?
mersonni (white
Wen
10-19.9
?
Mixed
?
Oon River,?
Hart (1947 17)?
Upper?
5?
33.6951 -1.1797?
2
?
?
21.5 27.0
??
26.3
(mode)
? Thornhill,
?
10?
19.9890 -0.6410
?
3?
-0.6851 29
?
28
??
27.7
Ontario?
15
?
31.9007 -1.0034
?
2?
?
30?
29.5
??
29.3
20?
27.0023 -0.81168
?
4?
-0.9606 31.5 30
??
29.3
25??
22.2209 -0.6211
?
1?
-0.9888 32.5 29.5
??
29.3
Lower
?
20?
2.5
25
?
........
?
6.0
(1152),14
.11 is assumed in this table that the acclimation temperature reported Is a true acclimation in the context of Brett
•
sillo
Correlation
bar
of median
coefficient
resistance
(perfect
times
M of
used
all data
for
points
calculating
to till
regression
regression
equation.
line=1.0).
d
=Incipient lethal temperature of Fry, et al,
(1946).81
x Salinity.
/ Log time in hours to
50%
mortality. Includes 2-3 hr. required for test bath to
reach
the test temperature.
From: National Academy of Sciences (1973). See pp. 410-419, 444-445, Appendix II-C.
Acclimation?
log time= a-l-h (temp.)
Species?
Stage/age?
Length?
Weight?
Sex
Location?
Reference
Extreme
Temp
?
Time?
a
?
h
?
N°
?
r°
Appendix II-C1411..,
THERMAL TABLES-Continued
Data limits
?
Lethal
(°C)?
1.050
?
thresholds
(°C)
upper lower
Upper?
2?
8 wks
?
16.5135 -0.6689?
4?
-0.9789 23.0 19.0
?
?
19,1
5?
4 wks?
10.2199 -0.3645
?
3?
-0.9264 24.0 20.0 ??
21.7
10
?
>2 wks?
12.4993 -0.4098
?
6?
-0.9134 28.0 24.0
??
24.2 -
20
?
2 wks?
11.2967 -0.5333?
8?
-0.9481 30.0 26.0 ??
26.2
25?
3 wks?
15.1204 -0.4493?
1
?
-0.9164 30.0 25.5
??
25.1(x)
Lower?
2?
8 wks
?
1.5?
0.3 ?
<0.3
5?
4 wks
?
1.0?
0.5??
<0.5 "-.-
10?
> 2 wks?
2.1355
?
0.3381
?
5?
0.9021?
3.0?
0.5 ??
3,0 7..;:
20
?
2 wks?
2.5090?
0.2685
?
6?
0.9637 4.5?
0.5
??
4.7
25
?
3 wks?
1.1154?
0.1652?
9?
0.9175 9.5
?
0.5
?
?
9.7
Upper?
5?
11 dae?
15.8243 -0.5831?
5?
-0.9095 26.0 22.0
??
22,2
10?
5 da?
9.0100 -0.2896?
6
?
-0.9516 30.0 23.0 ??
23.6
15?
5 da?
17.1908 -0.5101
?
4?
-0.9960 28.0 24.5
??
24.1
20?
5 da?
28.6392 -0.9451?
4?
-0.9692 :29.0 25.5
??
26.2 -.
25
?
5 da?
21.3511 -0.6594
?
5?
-0.9958 30.0 26.5
??
26.7
Caregonus astedii Juvenile?
?
Mixed
?
Pickerel?
Edsa II and
(deco)
?
Lake,.
?
Colby,
Washtenaw?
1910100
Co., Mich.
Coregonus hoyi?
Juvenile?
60.0 mm
??
Mixed
?
Lake Midi-
?
Edsall, Bottlers
(bloater)
?
(age 1)
?
5.0. 5.8
?
gun atf?
& Brown,
Kenosha,?
197040
Wisc.
Uppei
?
35?
(0
v
/oo)?
21.9021 -0.6211?
6?
-0.9183 43.0 40.5
35?
(5 0 /0)?
35.3415 -0.1855
?
6?
-0.9787 43.5 41.0
??
40.5
35?
(10 °too)?
30.0910 -0.6629?
6
?
-0.9950 43.5 41.5
35?
(20 0/iia)
?
30.0394 -0.6594
?
4?
-0.9982 43.5 41.5
Upper?
30?
700 hirs.h
35.0420 -0.8025
?
2
?
40.8 ?
(from 21.3 C)
Upper
?
25?
field &
?
47.1163 -1.3010?
3?
-0.9915 35.5 34.5
??
34.0
3-4 da
30?
38.0658 -0.9694
?
4?
-0.9921 38.0 36.5
??
36.0 ..,.
35?
31.5434 -0.7110
?
5?
-9.9642 39.0 31.0
?
?
36.50)
Lower
?
15?
10.1 •
30?
14.5
35?
20.0
Cyprinodon rade- Adult
?
?
Jefferson?
Strawn and
plus (sheeps-
?
County,
?
Dunn
head minnow)
?
Texas?
(196190)
Cyprinodon vane- Adult
?
?
Galveston?
Simmons
gatus variegatus
?
Island, Gal-?
(1911)"
(sheepshead
?
veston, Texas
minnow)
Dorosoma cepedi- Underyearling
?
?
Put-in-Bay,
?
Hart (1952)"
anum (gizzard
?
Ohio
shad)
Dorosoma cepedi- Underyearling
?
?
?
Knoxville,?
Hart (1952)44
anum (gizzard
?
Tenn.
shad)
Esox luaus?
Juvenile?
Minimum?
?
Maple, On-
?
Scott (1964)95
(Northern Pike)?
• 5.0 cm?
tario, Canada
Esox masquinongy Juvenile ?
Minimum?
?
Beadlike
?
Scott (1964)"
(Muskellunge)?
5.0 cm?
Hatchery
Ontario,
Canada
Esox hybrid?
Juvenile?
5.0 cm?
?
Maple, On-?
Scott (1964)00
(luclusx masqui-?
minimum
?
tario, Canada
nongy)
Fundulus chryso- Adult
?
?
Jefferson?
Strawn & Dunn
Ws (golden top-
?
County,?
(1961)"
minnow?
Texas
Fundulus diapha- Adult?
?
Halifax Co.
?
Garside and
nut (banded?
and Annapo- Jordan
killifish)?
fis Co., Nova (1968)44
Scotia
Fundulus orandis Adult
?
?
Jefferson?
Shwa &
(guff killifish)
?
County,?
Dunn
Texas
?
(1961)0
Upper?
25
?
32.1348 -0.8698
30
?
41.1030 -0.0541
35
?
33.2846 -0.8116
Upper?
25.0
?
11.3066 -0.4523
27.5
?
11.4439 -0.4490
30.0
?
11.0961 -0.4319
Upper?
25.0
?
18.1879 -0.5035
21.5
?
20.0811 -0.5283
30.0
?
18.9506 -0.4851
Upper?
35?
(0
0
/00)- 23.7214 -0.5219
35?
(5
v
/c0)- 21.2575 -0.4601
35?
mum-
21.8635 -0.4759
Upper
?
15?
(0 Vco)i ?
15?
(14 oho)
?
15?
(32
°/00 ?
Upper?
35?
(0 o
/oo)?
22.0809 -0.5119
35?
(5 Vol)?
21.6441 -0. 6220
35
?
(10 °/m)
?
24.9072 -0.5535
35?
(20 °AO
?
23.4251 -0.5169
2??
35.5 35.0
??
34.5
4?
-0.9991 38.0 36.5 ??
38.0
6?
-0.9896 39
?
36.5 ??
36.5
5
?
-0.9990 34.5 32.5
??
32.25
5
?
-0.9915 35.0
?
33.0
??
32.15
5?
-0.9911 35.5 33.5
??
33.25(1)-
5?
-0.9142 34.5 32.5
?
5?
-0.9911 35.0
?
33.0 ??
32.11
5?
---0.9972 35.5?
33.5 ??
33.29.
(5),'
32.5
37.75
33.25
(a)
3I.5
.
?
.
8?
-0.9182 42.0?
31.5 ?
. . . .. .•
1?
-0.9961 42.5?
39.5 ?
.. ....-
9?
-0.9926 43.0
?
39.0 ?
.... :. . ,
8
?
-0.9970 43.0 39.5 ?
Upper
?
25.0
?
18.6533 -0.4926
?
4?
-0. 9941
34.5 33.0
?
21.5
?
20.1834 -0.5460
?
5
?
-0.9995 35.0 33.0
?
30.0?
19.6126 -0.5032
?
5?
-0.9951 35.5 33.5
?
9
?
-0.9968 43.0 39.0 ?
-0.9969 43.5 40.0
?
8
?
-0.9905 43.5 40.0
?
Fundulus hetero- Adult
?
Halifax
?
Co.?
Garside and
clitus (mummic-?
and Annapo- Jordan
hog)?
lie Co., Nova
(1968)84
Scotia
It is assumed in this table that the acclimation temperature reported is 1
true acclimation in the context of Brett
(1952):14
Number of median resistance times used for calculating regression equation.
•
Correlation coefficient (perfect fit of all data points to the regression line=1.0).
d
=Incipient
lethal temperature of Fry, et al., (1946)."
•
Experimental fish were hatched from eggs obtained from adults from this location.
Upper
?
15?
(0 0/4 i
?
15?
(14 0/co)
15?
(32 °/w) ?
Experimental fish were reared from eggs taken from adults from this location.
These times after holding at 8 C for >1 mo.
h
Acclimated and tested at 10
v /ao salinity.
Tested in three salinities.
Tested at 3 levels of salinity.
412/ Appendix II-Freshwater Aquatic Life and Wildlife
THERMAL TABLES-Continued
Acclimation
Species
?
Stage/age?
Length?
Weight?
Sex
?
Location?
Reference?
Extreme ?
Temp.?Time
log time =a+b (temp.)
?
Data limits?
Lethal
(n)?
LD50?
thresholds
b?
Nb
?
r9?
(*C)
upper lower
Fundulus par-
?
Adult
?
6-7 cm
?
? Mixed?
Mission Ray, Doudoroff
?
Upper
?
14?
23.3781 -0.6439
?
4
?
-0.9845 34.0 32.0 ??
32.3
vipinnis (Cali-
?
Calif. (sea-?
(1945)
11
?20?
50.6021 -1.3451
?
11?
-0.9236 37.0?34.0 ??
34.4
fornia killifish)
?
water)
?
28?
24.5421 -0.5801?
7?
-0.9960 40.0 36.0 ??
36.5
(tested in seawater
?
Lower?
14?
2.1908?
1.0751?
3?
0.9449?
1.6?
0.4 ??
1.2
except as noted)
?
20?
2.1381?
0.2169?
6?
0.9469?
1.0?
2.0 ??
5.6
20?
2.5635
?
0.3481?
4?
0.8291?
4.0
?
2.0 ??
3.6
20?
(into 45%
?
2.6552?
0.4014
?
8
?
0.1348 4.0?
2.0 ??
3.8
sea water 1 day before
testing)
Fundulus
?
Adult
?
Jefferson??
Strewn and
?
Upper?
35
?
(0
0
/00)?
28.1418 -0.6304?
8?
-0.9141 43.0 39.0 ??
38.5
recess (bayou
?
County,?
Dunn?
35
?
(5 0
/0Q)?
29.3174 -0.6514?
1?
-0.9131 43.5 40.0 ?
killifish)
?
Texas?
(1967)79
?
35?
(100/00)
?
25.0890 -0.5477?
5?
-0.9956 43.5 41.5 ?
35?
(20 Voo)
?
30.4702 -0.6145
?
8?
-0.9849 43.5 40.0 ?
Fundulus similis Adult
?
?
Jefferson
?
Shawn and?
Upper?
35?
(0 "AO
22.9485 -0.5113
?
6?
-0.9892 43.0 40.5
(longnose killi-
?
County,?
Dunn?
35 -?
(5 0/0
?
25.6165 -0.5690
?
6
?
-0.9984 43.5 41.0
?
fish)?
Texas
?
(1967)11
?
35?
(10 Voo)
?
26.4675 -0.5863?
6?
-0.9925 43.5 41.0
?
35
?
(20.0/00)?
26.5612 -0.5819?
6?
-0.9953 43.0 40.5 ?
Gambusia Anis
.
Adult?
?
Mixed?
Knoxville,?
Hart (1952)14?
Upper?
25 ?
?
39.0004 -0.9771?
2?
39?
38?
37.0
antis (mosquito-
?
Tenn.?
30 ?
30.1523 -0.1143?
6?
-0.9938 40?
37.5 ??
37.0
fish)?
35?
23.8110 -0.5408
?
6?
-0.9978 4L5 39 ??
31.0(u)
Gambusia ethnic Adult
?
?
Jefferson Co., Strewn L
?
Upper?
35?
(0 ./
00)
.
22.4434 -0.5108?
5?
-0.9600 42.0 40.0 ?
(mosquitofish)
?
Texas
?
Dunn?
35?
(5 Voo)
?
23.1338 -0.5214
?
5
?
-0.9825 42.5 40.5 ?
(freshwater)
?
(1961)"?
35?
(10 cVeo)?
23.4911 -0.5304?
8?
-0.9852 42.5 40.0
35?
(20 Voo)?
22.1994 -0.5001?
6?
-0.9881 42.5 40.0 ?
Gamhusii affinis Adult
?
?
Jefferson Co., Strewn and?
Upper?
35?
(0 0/00). 11.6144 -0.3909
?
5?
-0.9822 42.5 40.5 ?
(mosquitolish)
?
Texas
?
Dunn'?
35
?
(5 0/
0 o)
?
18.9339 -0.4182?
5?
-0.9990 42.5 40.5 ?
(saltwater)?
(1961)19
?
35
?
(10 0
/0o)?
23.0784 -0.5165?
1?
-0.9982 42.5 39.5 ?
35?
(20 Voo)?
22.8663 -0.5124?
6?
-0.9951 42.5 40.0 ?
Gamhusia aMnis Adult?
? Mixed?
Welaka,?
Hart (1952)31?
Upper?
15?
32.4692 -0.8507?
3?
-0.9813 37?
35?35.5
holbrooki?
Florida
?
20?
38.3139 -0.9613
?
3?
-0.9843 38.5 37.5 ??
37.0
(mosquitofish)?
30?
31.4312 -0.1411
?
5
?
-0.9995 40
?
38
??
-?
37.0
35?
28.1212 -0.6564?
5?
-0.9909 40?
38.5 ??
31.0(u)
Lower?
15?
1.5
20?
5.5
35
??
14.5
Garmannia
?
. Adult?
?
Northern Gulf Heath (1961)11
?
Upper?
32?
21.7179 -0.5166?
3?
-0.9905 37.0 36.0 ?
chiquita (goby)
?
of California
Coast
Gasterosteus acu- Adult?
37 mm ave.?
0.50
g ave.?
Mixed?
Columbia?
Blahm and?
Upper?
19
lotus (three-
?
River near?
Parente
spine shale-
?
Prescott,?
(1910)01 un-
back)?
Oregon?
published
data
19.3491 -9.5940?
3?
-0.9993 32?
26 ??
25.8
Girdle nigricans Juvenile
?
7.1-8.0 cm?
? Mixed
?
Wall', Cali- Doudoroff
?
Upper?
12?
21.1217 -0.6339?
6
?
-0.9338 31.0 21.0 ??
28.7
(opaleye)
?
lornia (3314) (1942)"?
20?
19.2641 -0.5080?
7?
-0.9930 35.0?
31.0 ??
31.4
28
? 24.1213 -0.6140
?
4?
-0.9822 33.0 31.0
??
31.4
Lower?
12?
1.4851?
0.4886?
8?
0.955E?
5.0?
1.0?5.5
20?
?
-1.3818
?
0.6248
?
6?
0.9895 8.0
?
5.0 ??
8.5
21
?
?
-0.1238?
0.2614?
6?
0.9120 13.0?
6.0 ??
13.5
!datums
?
? Florida to On- Hart (1952)111?
Upper?
5? 14.6802 -0.4539
?
4?
-0.9182 29.5 28.0 ??
27.8
(Amicurus) nab-
?
brio (4 lo-?
10? 16.4227 -0.4842?
10?
-0.9526 31.5 29.5 ??
29.0
Moses (brown
?
cations) corn-?
15? 28.3281 -0.8239?
3?
-0.9881 33.0 32.5?31.0
bullhead)
?
bined
?
20?
23.9586 -0.6413?
11?
-0.9112 35.0?
32.5 ??
32.5
25
?
?
22.4910 -0.5132 12?
-0.9794 31.0 34.0 ??
33.8
30??
24.2203 -0.5917?
19?
-0.9938 38.5 35.5 ??
34.8
34?
19.3194 -0.4500?
5?
-0.9912 31.5 36.0
??
34.1
Lower
?
20?
0.5
25
?
4.0
30
?
6.8
I c
t
alurus ;uncle- Juvenile
?
?
Mixed?
Centerton,?
Allen &?
Upper?
26?
34.1119 -0.8816
?
13?
-0.9793 39.0?
36.6 ??
36.6
tos (channel
?
(44-57 da
?
Ark.?
Strewn?
30
? 32.1736 -0.1811
?
11?
-0.9510 40.6 31.4
??
37.8
catfish)
?
old)?
(hatchery)?
(1968) 72?34?
?
26.4204 -0.6149 20
?
-0.9638 42.0 38.0 ??
38.0
It is assumed in this table that the acclimation temperature reported is a true acclimation in the context of Brett
• (1952),74
N
umber of median resistance times used for calculating regression equation.
Correlation coefficient (perfect fit of all data points to the regression line= 1.0).
a
=Incipient lethal temperature of Fry, et al., (1946)."3
• Salinity.
53
Appendix II-C/413
413
THERMAL TABLES-Continued
Acclimation
?
log time=a+b (temp.)?
Data Omits?Lethal
Species
?
Stage/age?
Length?
Weight?
Sex?
Location?
Reference
?
Extreme ?
?
CC)
?
11350?
thresholds
Temp.
?Time?
a?
b?
Nb?
r°
?
CC)
upper lower
!Maus tonal- Juvenile
?
?
Joe Hogan
?
Allen 1,?
Upper?
25?
34.5554?
0.84154?
5?
-0.9746 37.5 35.5
??
35.5
tus (channel
?
(11.5 mo)?
State Fish?
Strewn
?
30?
17.7125 -0.4058?
4?
-0.934 40.0 37.5
??
37.0
catfish)
?
Hatchery,?
(1968)12
?35?
??
28.3031 -0.6554?
4?
-0.991/0 41.0 38.0 ??
38
Lonoke,
Arkansas
Ictalurus puncta- Adult
?
?
Mixed?
Welaka, Fla. Hart (1952)
44
?Upper
?
15?
34.7829 -1.0637
?
3
?
-0.9999 31.5 30.5 ??
30.4
tus (I. lacustris)
?
and Put-ln.
?20
?
39.4961 -1.1234?
4?
-0.9980 34.0?
33.0 ??
32.8
(channel catfish)?
Bay, Ohio?
25?
46.2155 -1.2899
?
5?
,
-0.9925 35.0 34.0
??
33.5
Lower
?
15
?
0.0
20?
0.0
25
?
0.0
Lepomis macro- Adult?
?
Mixed
?
Welaka,?
Hart (1952)
41
?Upper?
15?
25.2708 -0.7348
?
5?
-0.9946 33.0 31.0 ??
30:5
chirus purports-?
.
?
Florida?
20
?
28.0663 -0.7826
?
6?
-0.9971 34.5 32.5 ??
32.0
tens (bluegill)
?
.
?
25?
23.8133 -0.6320?
10?
-0.9150 36.0 33.0
??
33.0
30
?
25.1132 -0.6581?
5
?
-0.9965 38?
34.5 ??
33.8
Lower
?
15?
2.5
20?
5.0
25
?
7.5
30
??
11.0
Lepomis macro
.
?Adult?
?
Mixed?
Lake Mendota, Hart (1952)"
?
Upper?
20-23?
38.6241 -1.0581
?
4?
-0.8892 35.5 34.0 ?
chirus (bluegill)?
Wisconsin?
30?
??
30.1609 -0.7657
?
4
?
-0.9401 38.0 '36.0 ?
Lepomis megalotis Juvenile?
>12 mm?
?
Mixed?
Middle Fork, Neill, Strewn 1,
?
Upper?
25?
35.4953 -0.9331
?
14?
-0.9821 36.9 35.4 ??
35.6
(longear sunfish)?
White River, Dunn
?
30?
20.5581 -0.4918 22
?
-0.9625 39.0 36.5
??
36.8
Arkansas?
(1966)4?
35?
30.1245 -9.7257 43
?
-0.9664 41.5 37.3
??
31.5
Lepomis sym.?Adult
?
?
Jefferson Co., Strewn 1
?
Upper?
35?
(0 0/00
0
20.7487 -0.4686?
1?
-0.9747 42.0 39.0 ?
metricus (ban .
?Texas
?
Dunn
?
35?
(50/00)
?
23.5649 -0.5354
?
6?
-0.9975 42.0 39.0
?
lam sunfish)?
(1961)"
? 35?
(20
0
/00?
10.4421 -0.2243?
5?
-0.9813 41.5 39.5 ?
Lucania pun
?
Adult?
?
Jefferson Co., Strawn and
?
Upper?
35?
(0
om•?
21.2616 -0.4162
?
9?
-0.9844 42.5 38.5 ?
(rainwater ki=ll-?
Texas
?
Dunn?
35?
(5 0/00)?
24.3016 -0.5460
?
8
?
-0.9146 42.5 39.0
fish)
?
(1961)".?
35?
(100/0o)
?
24.3118 -0.5461
?
8?
-0.9904 42.5 39.0
35?
(20 0/0o)
?
21.1302
-0.4691
?
7-' -0.9940 42.5 39.5
Menidie menidla
?
8.3-9.2 cm?
4.3-5.2 gm?
Mixed?
New Jersey
?
Hot L West-
?
Upper?
1?
19,8801 -0.1391?
5?
-0.9398 24.0 20?
22.0
(common silver-?
(average?
(average?
(40°N)?
man (1906)/0
?14
?
18.7499 -0.6001?
6?
-0.9616 21.0 23.0 ??
25.0
side)?
for test?
for test?
21?
65.1350 -2.0387?
6?
-0.9626 32.0 28.0
??
30.4
groups)?
groups) •?
28
?
31.6032 -1.0582?
5?
-0.8112 34.0 30 ??
32.5
Lower?
7?
?
-9.8144?
8.9019?
5?
0.8274 2?
1?
1.5
14?
?
-1.2H4?
2.5597?
6 . 0.1594 5?
1?
2.0
21?
?
-1.4801?
L1484?
6?
0.9531 1
?
2
?
4.3
28?
?
-8.2366?
1.3586?
5?
0.9130 15?
7?
8.
1
Micropterus sal- 9-11 mo. age
?
?
Welaka,?
Hart (1952)48?
Upper?
20
?
35.5101 -1.0112
?
5?
-0.9787 34
?
32?
32
mcides Rod-?
Florida?
25?
19.9911 -0.5123?
8?
-0.9972 36.5 33
?
33
danus (large-?
30?
11.5645 -0.4200
?
8?
-0.9920 38
?
34.5 ??
33.1 (a):
mouth bass)?
Lower
?
20?
5.2
25?
7.0
30
??
10.5
Miuopterus sal .?
?
Put.in•Bay,?
Hart (1952)44?
Upper?
20?
50.6091 -1.4639?
2?
?
34?
33?
32.5
mo=des (large-
?
Ohio
?
25
?
26.3169 -0.6846
?
3?
-0.9913 36.5 35?
34.5 -
mouth bass)?
30
?
?
21.0213 -0.1150
?
4?
-0.9959 38.5 37
?
36.4(e)
Lower?
20?
5.5 *-..,.
30?
11.8
Micropterus sal- Under yearling
?
?
?
Knoxville,?
Hart (1952)
4
?Upper?
30?
36.0620 -0.9055
?
4?
-0.9788 38.5 37
?
36.4
moides (large-?
Tenn.
?
35
?
23.9185 -0.5632
?
6?
-0.9958 40?
37.5
??
35.4(4
mouth bass)
Micropterus sat-
?
?
?
Lake Men-
?
Hart (1952)
44
?Upper?
22?
34.3649 -0.9789
?
4?
-0.9189 33.1 32.0 ?
?
31.5
moides (large-
?
dole, Wis
.
?30?
35.2111 -0.9084
?
4
?
-0.9845 37.5?
35.5
?
. ..... ...:
mouth bass)?
cousin
..-
Mysis relicts
?
'Adult?
?
Mixed?
Trout Lake,?
Smith (1910)"?
Upper?
7.56
?
>1 wk?
6.1302 -0.1470
?
3?
0.9245 26?
16?
16
(Opposum?
Cook
shrimp)?
County,
Minnesota
• It is assumed In this table that the acclimation temperature reported is a true acclimaffon in the context of Brett
?
• Correlation coefficient (perfect fit of all data points to the regression line= 1.0).
(1952).74
?
d
=Incipient lethal temperature of Fry, et al., (1946).46
b
Number of median resistance times used for calculating regression equation.
?
• Salinity.
54
.414/ APPendix 1I-Freshwater Aquatic Life and Wildlife
THERMAL TABLES-Continued
Acclimation
log time=a+b (temp.)?
Data Omits?
Lethal
Species
Stage/age
Length?
Weight?
Sex?
Location Reference
?
Extreme ?
?
(°C)?
1.050
?
thresholds
Temp.
?Time?
b?
Ne?
r.?
(n)
upper lower
Hom
o
swat- Adult
?
>7 mm
tchensis
(o
pos
-sum shrimp)
trotemigonus
?
Adult
crysoleucas
(golden shiner)
Notropis alherl- Juvenile
noides (emerald (<1yr)
shiner)
? ?
Composite.?
Hart (1952)"
Upper
10
of 1. Welaka,
Fla. 2. Put-
15
20
in
,
Bay, Ohio
25
3. Algonquin
30
Park, On-
tario
Lower
15
20
25
30
? ?
0-1.9 g.mode?
Mixed
Chippewa?
Hart
(1941)"
Upper..
5
Creek, Wel-
land, Ontario
10
15
20
25
Lower
15
20
25
? ?
Toronto, On-?
Hart (1952)"
Upper
10
tarie
15
20
25(win-??
ter)
? Mixed?
Sacramento- Hair (1971)16
San Joaquin
delta, Cali-
fornia
Upper?
10.3.?
? 13 (48)
11.0?
?
12.5(49)
15.1?
?
13.8(49)
18.3?
?
16.1(48)
19.0?
?
14.0(48)
19.0? 9.4694 -0.2150?
2
?
?
24.2-25.41
21.7?
?
11.0(40 ?
22.0?
?
77.5(48)
?
22.4?
?
16.0(48)
42.7095 -1.3501
?
3?
-0.9998 30.5 29.5 ??
29.5
30.2861 -0.8933
?
4?
-0.9844 32.5 31.0 ??
30.5
31.0215 -0.1722 15
?
-0.9869 34.5 32.0
??
32.0
34.2505 -0.9226?
9?
-0.9665 36.0 34
??
33.5
26.3829 -0.6615 10
?
-0.9940 37.5 35
??
34.5
1.5
4.0
1.0
11.2
20.9532 -0.1959?
1?
-0.9519 24.5 23.5 ??
23.3
36.5023 -1.2736?
2?
?
27.5 27.0 ??
26.1
41.4849 -1.5441?
3?
-0.9803 30.5 29.5 ??
28.9
33.4114 -0.9858?
3?
-0.9805 32.5 31.5 ??
30.7
26.7046 -0.7337?
6?
-0.9153 34.0 31.5 ??
30.7
1.6
5.2
8.0
Nohopis cornutus Adult?
4.0-5.9 ?
g
?
Mixed
(common
?
(mostly 2 yr)?
(mode)
shiner)
Notropis cornutus Adult
??
? Knoxville,
•
(common shiner)
?
Tenn.
losorhynchos
?
Juvenile fresh . 3.81+0.29
?
0.30+0.151 Mixed
?
Dungeness,
•
• gorbuscha (pink
?
water fry
?
cm
?
Wash.
salmon)
?
(3.8
mo.)
?
(hatchery)
1??
29.0 29.0 ??
29.0
45.4331 -1.3979
?
2??
31.5 31.0
??
30.5
34.5324 -1.0116?
4?
-0.9560 33.0 31.5 ??
31.0
24.9621 -0.6878?
5?
-0.9915 34.0 32.0
??
31.0
25?
28.5059 -0.7741?
8?
-0.9913 35.5 32.0 ??
31.0
30? 28.1261 -0.1316?
6?
-0.9946 36.5 34.0 ??
31.0(u) -
Upper?
5
??
26.7
10?
40.7738 -1.3522?
3?
-0.9729 30.0 29.0 ??
28.6
15?
45.0972 -1.3814
?
3?
-0.9999 32.0 31.0 ??
30.3
20? 34.5324 -1.0116?
4?
-0.9560 33.0 31.5
??
31.0
25?
24.9620 -0.6878?
5?
-0.9915 34.0 32.0 ??
31.0
Lower?
20?
3.7
25?
7.8
Hart-(1952)"
?
Upper?
25?
25.5152 -0.6794?
6?
-0.9938 35.5 33.0
??
33.0
30? 24.9660 -0.6297 10?
-0.9979 WO 34.5 ??
33.5(a)
Brett (1952) 74
?Upper?
5?
11.1821 -0.4215?
4?
-0.9513 24.0 22.0 ?
21.3+0.3
10?
11.9021 -0.3865?
8?
-0.9840 26.5 23.0 ?
22.5+0.3
15?
12.1937 -0.4074?
8?
-0.9984 27.0 23.5 ?
23.1+0.3
20
?
16.2444 -0.4014?
1?
-0.9681 27.5 24.0 ?
23.9+0.6
24?
14.1111 -0.4459?
6?
-0.9690 21.5 24.5 ??
23.9
Don River,?
Hart (1947)10
Thornhill,
Ontario
Nolropis comutus Adult
(common shiner)
Olt
Cothynchus
?
Juvenile fresh- 5.44+0.89?
1.62+1.031
?
Mixed
. kola (chum
?
water fry
?
cm
salmon)
?
(4.9 mo.)
Nile Creek,?
Brett (1952)74
B
.C.
(hatchery)
Octotnynchus
?
Juvenile
?
? Big Creek?
Blahm and
Hatchery,
?
Parente
Hoodsport,?
(1970)100
Wash.h
?unpublished
data
(11.52),rs
. 1111 e
ssumed in this table that the ocarnatiOn temperature reported is a true acclimation in the context of Brett
1.
•
H
umber of median resistance times used foe calculatingg regression
equeffoo
-
?
-
a:I.144c coe
fficient (perfectregression lins=1.0).
ot
elent MIMI temperature of Fry, et al., (1946)."
•e0 temperatures estimated from a graph.
kite (chum
salmon)
Upper?
5?
14.3829 -0.5320?
4?
-0.9839 24.0 22.0 ??
21.8
10?
14.1773 -0.4166
?
9?
-0.2665 26.5 22.5 ??
22.6
15?
15.9911 -0.5252
?
8?
-0.9070 27.0 23.0 ?
23.1+0.4
20?
16.1894 -0.5168?
9?
-0.9750 27.5 23.5 ??
23.7
23?
15.3825 -0.4121?
4?
-0.9652 21.0 24.0
?
23.8+0.4
Lower
?
5
10?
1?
0.5
15?
5
?
4.1
20?
7?
6.5
23?
? 8?
7.3
Upper?
9?
10%i?
16.9245 -0.5995?
6?
-0.9927 29?
11?22.0
50%?
15.9272 -0.5515?
4?
-0.9912 29?
17?23.2
90%?
16.1763 -0.5181?
4?
-0.9995 29?
11
?
23.6
For maximum of
48
hr exposure. The lower temperature is uncorrected for heavy mortality of control animals at
"acclimation" temperatures above about 21.6.
The author concluded that there were no geographic differences. The Welaka, Florida subspecies was itches',
the others N.c. :stratus, based on morphology.
Tasted in Columbia River Water at Prescott, Oregon.
Mortality Value.
55
Appendix II-C/415
THERMAL TABLES-Continued
Species?
Stage/age
?
Length
?
Weight?
Sex
Oncorhynchus?
Juvenile fresh- 4.18±0.6?
1. 37±0. 62g Mixed
Kisutch (coho?
water fry?
cm
salmon)
?
(5.2 mo.)
Acclimation?
log time= 3+6 (temp.)
?
Data limits?
Lethal
Location
?
Reference
?
Extreme ?
?
(°C)
?
LG150
?
threshold,'
Tempo?
Time?
b
?
(°C)
upper lower
Nile Creek,?
Brett (1952)
74
?Upper?
5
??
21.3050 -0.1970
?
2?
?
24.0 23.0
B.C.
?
10?
19.5121 -0.6820
?
4?
-0.9841 26.0 24.5
(hatchery)
?
15?
20.4066 -0.6858?
6?
- 0.9681 21.0 24.5
20
?
20.4022 -0.6713
?
4?
-0.9985 27.5 25.5
23
?
18.9136 -0.6013
?
5?
-0.9956 21.5 25.0
Lower?
5?
0.2. •
10
??
1?
1.1
15
??
3?
3.5
20
??
5?
4.5
23
??
7?
1.0 ??
6.4
22.9±0.3
23.1
Onc
24.3±6.3
Is
25.0±0.2
(c
25.0±0.2
sa
Oncorhynchus
?
Juvenile
?
?
Kalama Falls, Blahm &
kisutch (calm
?
Wash.?
McConnell
salmon) .
?
(hatchery).?
(1910)00
unpublished
data
Oncorhynchus
?
Adult?
a 510 rem?
a 2500
g
ave. Mixed
kisutch (coho?
ave.
salmon)
Upper?
10?
(1 0%)I
?15. 4616 -0.5522
?
6?
- 0.8533 29
?
1.1 ??
23.2
(50%)?
11 4136 -0.6410
?
6?
-0.9105 29
?
11.0
?
23.5
(90%)
?
15.9026 -0.5423
?
4
?
-0.9110 29
?
11.6 ??
23.1
140?
(10%)?
8.5301 -0.2969
10
?
-0.9063 29
?
14.0 ??
14.0
(50%)
?
8.5195 -0.2433 10
?
-0.8483 29?
0.14 ??
17.0
(90%)
??
22.0?
i
Columbia?
Coutant?
Upper?
174?
5.5068 -0.1610
?
5?
-0.9167 30?
26
.
?
1 ,
River at?
(1910)71
Priest Rap-
ids Dam
Onto
Ish
(Ct
Spf.
ti
Issaquah,?
Brett (1952)
74
?Upper
?
5?
11.1881 - 0. 6623
?
4?
-0.9383 24.0
?
22.5 ?
22.2±0.3
Wash.
?
10?
14.1319 -0.4988
?
8?
-6.9833 26.5 23.5
?
23.4±0.7
(hatchery)?
15?
15.8199 -0.5210
?
1?
-0.9126 27.5
?
24.5
?
24.4±0.3
20
?
19.3121 -0.6318
?
5?
-0.5602 21.5
?
24.5 ?
24.1±0.3
23
?
20.0020 -0.6496
?
4?
-0.9981 26.5 24.5
?
24.8±0.1
Lower?
5?
0?
0?
0 ,.
10?
4?
0
?
3.1
15
?
5?
0? 4.1 '
20
?
5?
0? 4.1.
23
?
7?
1.0
?6.1
National Fish McConnell I?
Upper?
10?
10%)?
18.4111 - 0.6458
?
6?
-0.9611 29?
11
?
21.5
Hatchery'?
Blahm?
50%
?
18.5833 -0.6431
?
6?
-0.9750 29
?
11
?
22.5
Leaven-?
(1910)107?
90%?
20.6289 -0.1166?
6?
-0.9553 29
?
11
?
23.0
worth,
?
unpublished?
20?
10%?
11.5221 -0.5861
?
6?
-0.9739 29?
21
?
23.5
Wash.?
data
?
50%?
16.1328 - 0.5413
?
6?
-0.9552 29
?
21
?
23.5
. 90%?
15.1823 -0.5061
?
6?
-0.9539 29
?
21
?
23.5
Oncorhynchus
?
Juvenile fresh- 4.49±0.84?
0.81±0.45g Mixed
nerka (sockeye
?
water fry
?
cm
salmon)?
(4.7 me)
Oncorhynchus
?
Juvenile
?
61 mm a
y
e.
?? Mixed
nerka (sockeye
?
(under
salmon)
?
yearling)
Oncor.
tsha
(chU
Oncorh
!Shay
(chin
fall n
4
Oncorhynchus
?
Juvenile?
100-105 mm ?
Mixed
nerka (sockeye
?
(yearling)
?
are tor test
salmon)
?
groups
Oncorhynchus?
Juvenile fresh- 4. 44±0. 40
?
1. 03±0. 21g?
Mixed
tshiwylscha?
water fry
?
cm
(Chinook?
(3.6 mo.)
salmon)
National Fish McConnell &?
Upper?
10 1°C?
(16%))?
6.4771 -0.2118?
4?
-0.9881 32
?
14
..
13..
.
:5
"
'''
I --
1; P
ura
il
(Clki
nifI
nGic
worth,
Hatchery
?
Blahm
Wash.'
Leaven?
(1970)103
unpublished
data?
per day rise
to anal. temp.
12"?
(10%)
?
13.2412 -0.4415?
4?
-0.5955 29?
11
15.5"?
(10%)?
12.1763 -0.4004
?
5?
-0.5443 32
?
11
(50%)?
9.0438 -O. 2922?
4?
-0.9392 32
?
14
(90%)?
9.0628 -0.2859
?
4?
-0.9534 32?
14
(50%)
?
18.1322 -0.6118
?
4?
-0.9598 29?
11
(90%)
?
11.5421 -0.5900
?
4?
-0.9533 29?
17
(50%)
?
13.6666 -0.4432
?
5
?
-0.9120 32
?
17
(90%)?
12.1165 -0.4057
?
4
?
-0.9748 32
?
11
?
......... 'tt.
n.5
fi
?
(Yam
',ma
rl
?
.
.
2.3:5
.
.
..1 Ono
tthoi
rhwy
Dungeness,?
Breit (1952)7 1
?Upper?
5
Wash.
(hatchery)
Lower?
10
17 "?
(10%)?
11.4210 -0.6114
?
5?
-0.9549 29
?
20
10
20
24
15
15
20
23
?
(50%)?
11.2432 -0.5685
?
4?
-0.9450 29?
20
(90%)
?
11.2393 -0.5169
?
4?
-0.9364 29?
20
16.4595 -0.5515?
5?
-0.9996 26.5 24.5
?
16.4454 -0.5364?
4?
-0.9906 21.0 25.5 ?
22.9065 -0.1611
?
1?
-0.9850 21.5 25.0
?
18.9940 -0.5992
?
9?
-0.9923 21
31 ... 050
2500..05
?
9.3155 -0.3101
?
6
?
-0.9841 25.0 22.5 ?
5.0?
0.5 ?
3.0
?
1.0 ?
22355
4...3
014
101-.111i
;;
25.
14,
.
Poomyz
.....
•-
..
.,1
,
?•11155
.........
'?
(pilau
13.5
21.5?
"
il 4-
.?
merino
7.5 f"
?
ii
.1
1 .
I; 41'ed)
(and may have included a few fish from other upstream sources). River water was supersaturated with 00
,,
'Numb
0 14 C-acclimated fish were collected from the Columbia River 4-6
who following release from
the
0
:' (1:5:)
11P:re'al
and 14-C fish showed signs of gas-bubble disease during tests.
b
River temp. during fall migration.
i
Tested in Columbia River water at Prescott, Oregon.
i Per cent mortalities.
.
It is assumed in this table that the acclimation temperature reported is a true acclimation in the context of Brett
(1952).74
b
Number of median resistance times used I or calculating regression equation.
•
Correlation coefficient (perfect fit of all data points to the regression line = 1.0).
d =
Incipient lethal temperature of Fry, et al, (1946).47
•
10 C-acclimated fish came directly from the hatchery.
Data were presented allowing calculation of 10% and 90% mortality.
416/ Appendix II-Freshwater Aquatic Life and Wildlife
THERMAL TABLES-Continued
Species
?
Stage/age?
Length?
Weight?
Sex
Oncorhynchus
?
Juvenile
Dilly/Moho
(chMook
Almon)
Oncorhynchus
Juvenile?
14 mm are.?
6.3g are.?
Mixed
tsluivytscha
(Chinook salmon
'spring run)
Oncorhynchus
?
Juvenile
?
40 mm. are.?
?
Mixed
tshawytscha
(chinook salmon)
Location?
Reference?
Extreme
Columbia?
Snyder &?
Upper
River at?
Blahm
Prescott,?
(1970)105
Oregon?
unpublished
data
Little White?
Blahm &
Salmon,?
McConnell
River?
(1910)000
Hatchery,
?
unpublished
Cook,
?
data
. Washington
Eggs from?
Snyder &
Seattle,?
Blahm
Wash.?
(1910)005
raised from
?
unpublished
yoIk•sac?
data
stagqin
Columbia
Rivet water
at Prescott,
Oregon
tog time-=a+b (temp.)
Data limits
(°C)?
1050?
threshold,'
Lethal
b
(°C)
Nb
r.
upper?lower
16.8109 -0.5181
3
-0.9998
29
25
24.5
18.9110
-0.6621
5
-0.9918 29
23
22.9
17.0278 -0.5845
3
-0.9997
29
25
24.5
15.7101 -0.5403
8
-0.9255 29
20
23.5
15.1583
-0.5312
8
6
-0.9439
29
20
20.5
15.2525
-0.5130
8
-0.9360 29
20
23.5
18.2574 -0.6149
5
0
-0.9821
29
23
20.5
12.4058 -0.3974
6
-0.9608 32
17
20.0
10.1410 -0.3218
1
-0.9496
32
17
19.5
12.1368
-0.4040
6
-0.9753 32
11
23.0
13.3115 -0.4240
11
-0.9550 30
20
20.5
11.5122 -0.3745
12
-0.9413 30
20
20.0
14.2456 -0.4434
10
-0.9620
30
20
23.5
13.3696
-0.4691
4
-0.9504 29
17
23.0
14:6268
-0.5066
4
-0.9843
29'
17
23.5
19.2211 -0.6679
4
-0.9295 29
17
23.8
22.6664 -0.7797
4
-0.9747 19
21
23.8
21.3981 -0.1253
3
-0.9579 29
21
24.1
20.5294 -0.7024
3
-0.9463 29
21
24.8
13.5019 -0.4874
4
-0.9845 29
8
20
8.9126 -0.3198
6
-0.9618 29
8
13.5
10.6491 -0.3771
6
-0.9991 29
8
2
Acclimation
Temp.
?Time
(10%/)
(90%)
10 g
(10%)
(90
%)
12
13
(1
0
%)
(90%)
18g
(10%)
(90
030
)
Upper?
11?
2-3-wks
10%,
50%
90%
20 1C/day rise
from IOC
10%
50%
90%
Upper?
4
(10%)!
(90%)!
39-124 mm??
Mixed
averages
for various
test groups
Oncorhynchus
?
Juvenile?
90.6 mm ave. 7.8 g
ave.?
Mixed
tshawytscha
.--
,
•
(chinook salmon
:fall run)
Oncorhynchus
?
"Jacks"
?
2500 mm are. 2000 g. ave.
?
Males
tshawylscha
?
1-2 yrs old
(Chinook
salmon)
Little White
?
Blahm &
Salmon?
McConnell
Riverhatch•
?
(1970)000
ery, Cook,?
unpublished
Washington
?
data
- -
Columbia?
Coutant
River at?
(1970)75
Grand Rapids
Dam
Upper
?
11?
2-3 wks
107
04
?18.61189 -0.6569?
5?
-0.9618 29
?
17
50%?
20.5471 -0.7147
?
4?
-0.9283 29?
17
90%?
20.8560 -0.7231?
4?
-0.9245 29?
11
Upper?
20?
1C/day rise
from 10C
10%?
21.6756 -0.7438?
4?
-0.9550 29
?
21
50%?
22.2124 -0.7526?
4?
-0.9738 29?
21
90%
?
20.5162 -0.6860?
3?
-0.9475 29?
21
Upper?
170? 13.2502 -0.4121?
4?
-0.8206 30?
26
19 1
?9.4683 -0.2504?
4?
-0.9952 26?
22
23.5
24.2
24.5
24.5
24.5
24.5
22
41
:1•1
g
.,P51/0m7lon
?
Prolarree
m
erino: (su
1 •
?
.?
.
l
ocked)
Puree thrums Juvenile
?
49 mm ave.
?
1.2 g ave.?
Mixed?
Columbia?
Blahm and
River near?
Parente
Prescott;
?
(1910)101
Ore.?
unpublished
data
t
,.Pirca Onerous Adult (4 yr
?
? 8.0-9.9 g
?
Mixed?
Black Creek, Hart (1947)or
• G
em Perch)
?
mode)
?
mode?
Lake Sim-
coe, Ontario
(yellow perch)
? Great Lakes McCauley
(1963)"
Upper?
19?
field plus?
15.3601 -0.4126?
2?
?
38?
32
4 da.
Upper?
5 ??
7.0095 -0.2214?
9?
-0.9904 26.5 22.0 ??
21.3
11?
17.6536 -0.6021
?
2??
26.5?
26.0 ??
25.0
15?
12.4149 -0.3641?
5?
-0.9994 30.5 28.5 ??
27.7
25?
21.2118 -0.5909?
6?
-0.9698 33.0 30.0 ??
29.7
Lower?
25?
3.1
Upper?
15 and 21 . ??
17.5642 -0.4680 18
?
-0.9683 34
?
29
?
28.5
•
01
'
Un
(1552).7t
assumed in this table that the acclimation temperature reported is a true acclimation in the context of Brett
11
/9
e Nu
!Correlation
siricipient
mber Of
co
lethal
median
efficient
t
emperature
resistance
(perfect Ot
of
times
Fry,
of all
used
et
data
aL,
for
points
(1946)-u
calculating
to the
regression
regression
equation.
One= 1.0).
5h
it♦
testerl shortly alter capture by beach seine.
• 7414?
available
i
on calculation of 10% and 90% mortality of June lest groups.
These were likely synergistic effects of high N2 supersaturation in these tests.
A Excluding apparent long-term secondary mortality.
Data were available tor 10% and 90% mortality as well as 50%,
Data also available on 10% and 90% mortality.
k
Data available for 10% and 90% mortality as well a; 50%.
0
River temperatures during fall migrations two different years.
No difference was shown so data are lumped.
57
Appendix II-C/417
THERMAL TABLES-Continued
Acclimafion?log time= e+b (temp.)
?
Data Omits
?
Lethal
Reference
?
Extreme
?
?
(°C)?
LD50
?
threshold
Temp.?Time
?b?
446
?
r•
?
(°C)
upper lower
Species?
Stage/age?
Length?
Weight?
Sex?
Location
Pimephiles?
Adult (mostly
? mostly 0-2 g Mixed?
Etobicoke Cr, Hart (1947)40
(Hyborhynchus) 1 yr)?
Ontario
notatus (blunt.
nose minnow) •
Pimephales?
Adult (1 yr)??
2.0-3.9 g?
Mixed
?
Don River,?
Hart (1947)47
ptomain (fat-
?
mode?
Thornhill,
head minnow) ..?
Ontario.
Poecilla latlinne Adult
?
?
Jefferson Co, Strewn and
(Sall molly)
?
Texas?
Dunn
(1967)gg
Pontoporeia ells Adult?
?
Mixed?
Lake Superior Smith (1971)104
near Two
?
unpublished
Harbors,?
data
Minn.
Pstudopleuro-
?
? 6.0-7.1 cm
?
3.4-4.2 g?
Mixed?
New Jersey?
Hoff L West-
nectes amen.
?
(averages?
(averages? (40°44)?
man (1966)14
anus (winter?
for test .?
for test
flounder)
?
groups)?
groups)
Rhinichthys
?
Adult?
?
Knoxville,?
Hart (1952)"
atratulus
?
Term.
(blacknose dace)
Rhinichlhys?
Adult (7)?
?
Toronto,?
Hart (1952)44
atratulus (black.?
Ontario
nose dace)
Rbinichthys?
Adult?
2.0-3.9??
Mixed?
Don River,
?
Hart (1947)47
atratulus (Black.
?
(mode)?
Thornhill,
nose dace)
?
Ontario
Upper?
5
?
24.6411 -0.1602?
2??
27.0 26.5
??
26.0
10?
...
??
55.8357 -1.1588?
2??
29.5 29.0 ??
21.3
15
?
?
28.0371 -0.1337?
3?
-0.9974 32.0 31.0 ??
30.6
20
?
?
34.3240 -0.9982?
4?
-0.9329 34.0 32.5 ??
31.1
25? 50.1212 -1.4181?
3?
-0.9450 35.0 ILO ??
33.3
Lower
?
15
??
10
20?
4.2
25
?
7.5
Upper?
10
?
60.7712 -2.0000?
2??
30.0 29.5 ?
28.2
20?
6.9970 -0.1560?
4?
-0.7448 33.0 21.5 ??
31.7
30?
?
41.3696 -1.1317?
5?
-0.9610 36.0 34.0?33.2
Lower?
20?
1.5
30?
10.5
•
Upper?
35?
(0
0
/0),
27.4296 -0.6279?
6?
-0.9902 12.5 31.5
?
35?
, (5 0/00)?
25.6936 -0.5153?
6?
-0.9835 42.5 39.0
35?
(10
0
/00)?
21.8108 -0.6535?
7?
-0.9949 42.0 39.0 ?
35?
(29 0
/4
?
27.1918 -0.6146
?
3
?
-0.9191 42.5 39.5
?
Upper?
8
?
9.1790 -0.5017
?
2??
12?
10.1 ??
10.5
9?
? 10.4
(30 de)
Upper?
1? 21.2916 -1.1405?
4?
-0.9852 21.0 20.0
?
22.0
14
?
24.3020 -0.8762
?
6
?
-0.9507 26.0 23.0
?
23.7
21
?
49.0231 -1.6915?
5?
-0.9231 29.0 26.0
?
27.0
28
?
60.8010 -1.9610?
4?
-0.9181 30.0 29.0
?
29.1
Lower
?
1?
1.0?
1.0
?
1.0
14?
2.0
?
1.0
?
1.0
21
?
2.4924
?
0.1165?
3?
0.7816 6.0?
1.0
?
14
28
?
2.2145?
0.2344?
3?
0.9970 1.0?
4.0
?
6.0
Upper?
20? 21.2115 -0.5958
?
1
?
-0.9935 33
?
30
?
29.3
25
?
19.6451 -0.5224 10?
-0.9979 35?
30.5 ??
29.3
28
?
21.3360 -0.5651?
7?
-0.9946 35.5 32.5 ??
29.3
Upper
?
5?
?
27?
27?
27(1 hr)
15? 19.8151 -0.5111?
4?
-0.9632 31.5 30.0 ??
29.3
20?
24.5749 -0.1061?
7?
-0.9926 33?
30.0 ??
29.3
25?
20.1840 -0.5389?
I?
-0.9961 35
?
32.0
??
29.3
Upper?
5?
17.1811 -2.1959?
2?
?
21.5 27.0 ??
26.5
10? 49.1469 -1.6021
?
3?
-0.8521 30.5 29.5 ??
28.1
15?
19.6915 -0.5134
?
4
?
-0.9571 31.5 30.0 ??
29.6
20
? 26.5952 -0.7719?
II?
-0.9891 33.5 29.5 ??
29.3
25? 23.5165 -0.6629
?
9?
-0.9937 34.0 30.0
??
29.3
Lower?
20?
2.2
25?
5.0
Alabaster &
?
Upper
?
111f
?
11.4654 -0.5801?
5?
-0.9787 29.6 26.3
??
26.5
Welcomme
?
11•? 13.6531 -0.4264?
5?
-0.9742 29.1 26.3 ??
26.5
(1962)7°
Craigie, D.E.
?
Raised in soft water
(1963)70?
Upper?
20 (tested in soft
water)?
14.6405 -0.4470?
3
?
-0.9787 29?
21
20 (tested in hard
water)
?
15.0392 -0.4561?
3?
-0.9911 29?
27
Raised In hard water
20 (tasted in soft
water)
?
15.1473 -0.4683?
3?
-0.9781 29?
27
10 (tested in hard
water)
?
12.8718 -0.3831?
3?
-0.9841 29?
27
Salmo gairdnerii Juvenile
?
4.5-+0.4 cm ?
Mixed?
Britain
(Rainbow trout)
Salmo gairdneril Yearling
?
? East end of
(rainbow trout)?
Lake
Superior
Selma gairdnerii Juvenile?
9.4+6.0 ern ?
Mixed?
London,?
Alabaster &
(rainbow trout)
?
and 15.5+
?
England?
Downing
1.1 cm?
(Hatchery)?
(1966)44
•
It is assumed in this table that the acclimation temperature reported is a true acclimation in the context of Brett
(1952),74
Number of median resistance times used for calculating regression equation.
• Correlation coalfkient (perfect fit of all data points to the regression lint-1.0).
d
..Insicrisnt lethal temperature of Fry, at al., (1946).0
Upper?
15?
15.6500 -0.500?
24 ?
20?
19.6250 -0.6250?2 ?
• Salinity.
Diuolved oxygen Cone.7.4 mg/I.
g
Dissolved oxygen Conc. 3.8 mg/t.
4
See note (under Salmo sale') about Alabaster 1967.44
4
18/Appendix II-Freshwater Aquatic Life and Wildlife
THERMAL TABLES-Continued
Acclimation
?
log Ame=a-i-b (tamp.)
Location?
Referenu
?
Extreme
Temp.
?Time
?
b
upper lower
Shp/age
?
Length?
Weight
?
Six
Data limits
?
Lethal
CC)?
LD50?
threshold,'
CC)
Columbia?
Coutant?
Upper
River at?
(19707'4
Priest
Rapids Darn
?
Mixed?
River Axe,?
Alabaster?
Upper
Devon,?
(1461)••
England
goo
gairrinerii Adult
?
2650 mm?
4000
g ave.
?
Mixed
(ansdromous)
(31881hlid
trout)
Salmo
slat
Smofts
(1-2?
About 16 cm
(Atlantic salmon) yrs)
19.
?10.9677 -0.3329?
7?
-0.1910 29
?
21
9.2 (field)?
43.6661 -1.6667?
2r ??
(1)
?
(1)
9.3"?
23.7273 -0.9091?
2
10.9"?126.5000 -5.000
Tested in 30%
sender
E2 (fled)?
44.6667 -1.6867?
2 ?
Tested in 100% sea-
water
9.2 (Add)?
14.7361 -0.5263?
2 ?
Acclimated 7 hr in sea-
water; toted in
sea-
water
9.2 (field)
?
36.9999 -1.4216?
2
21
Salmo pier
?
Newly hatched
(Atlantic salmon) nine
Salmo Mar
?
30 da after
. (Atlantic salmon) hatching
?
Mixed?
Cullercoats,?
Fshei
(1350)
1
?Upper
North
Shields,
England
(hatchery)
?
Mixed?
Cullucoats,?
Sishal (1460) •
?Upper
North
Shields,
England
(hatchery)
6 (brought up to?
13.59?
-0.4217?
1?
-0.9671 21.0 20.0 ??
22.0
test temp. in
6 hours)
5
?
1.9631 -0.2177?
4?
-0.9791 25.0 22
?
22.2
10
?
15.7210 -0.5396?
3?
-0.1619 26.0 22
?
23.3
20
?
11.5471 -0.3406?
3?
-0.9143 21.0 22
?
23.5
Upper
Upper
Upper
Upper
?
Mixed?
River Axe,?
Alabaster
Devon,?
(1967)8
England
11.71.5 cm
?
Mixed?
River North Alabaster
Esk, Scotland (1967)•
14.161.3 cm ?
Mixed
Selma Mitts?
Newly hatched
?
?
Mixed
(brown trout)?
fry
9.3 (field)
?
33.3150 -1.2500
?
2' ?
10.9 (field)?
21.0000 -1.0000
?
2
11.7?25.9091 -0.9091?
2' ?
16.7?14.5909 -0.4545?
Ig ?
6 (raised to test
temp. over 6 hr
period)
?
12.7756 -0.4010
?
6?
-0.9747 21.0 20.1
??
22.0
Salmo sitar
?
Parr
(1
yr)?
10 cm an.
(Atlantic salmon)
Salm° olar?
Smalls (1-2
(Atlantic salmon) yrs)
Satrap
Mar?
Smells (1-2
(Atlantic salmon) yrs)
River Sewn Alabaster
Glouti
g
iil. (19617"
England
Cullernats,?
BONI (1960)11
North
Shields,
England
(hatchery)
Sxlmo truth
?
30 da after
?
?
Mixed
(Drown trout,
?
hatching
noun)
Cullercoats,?
Ilshai (1960)71
North
?
•
Shields,
England
(hatchery)
Upper?
5
??
15.2544 -0.5219?
4?
-0.1713 25.0 22.0 ??
22.2
10
??
23.5131 -0.1406?
3?
-0.9702 26.0 22.0 ??
23.4
20
??
14.1971 -0.4855?
3?
-0.9797 28.0 22.0 ??
21.9
Salmo butts
?
Juvenile?
10.160.1 cm ?
Mixed?
London,
?
Alabaster &
(brown trout,
?
7.4±4.5
?
England?
Downing
noun)?
cm?
(hatchery)
?
(1066)•
Selma tram
?
Smalls (2 yr.) About 21 cm
?
Mixed?
River Axe,?
Alabaster
(brown trout,
?
an.?
Devon,?
(1967)14
noun)?
England
Unitas lone- Juvenile
oak (kook
?
?
Piusant
?McCauley
Mount?
(1950)"
trout) Hatchery,
Wayne Co.,
Penna. and
Chatsworth
Hatchery,
Ontario",
0152),74
•
It in assumed in this table lint the acclimation temperature reported it a true acclimation in the context of Intl
4
•
Hu
etu
l
nter
bsoent
ddion
of median
coefficient
lethal temperature
resistance
(perfect
limas
of
At
Fry,
of
used
all
et
den
al.,
for calculating
points
(1944)."to
the
regression
regression
equation.
line-10).
Upper?
6?
?
36.1415 -1.4211?
2' ?
15?
21.5114 -0.7143?
2
20??
17.6667 -0.5556?
2 ?
Uppe?
9.3 (field)
?
11.4667 -0.6661?
2' ?
10.9"?33.0000 -1.2500?
2 ?
Upper
?
10
?
?
17.5260 -0.6033?
6 -0.9254 25.5 2(5
?
20??
20.2457 -0.6671
?
7 -0.9723 27.0 25.0
?
• Rim
temp. during fall migritiort
I
Alabaster fitted by eye,
a
straight line to median death times plotted on semllog paper (log time), thin reported
only
the
100 and 1000 min intercepts. Than intercepts
III
the basis for the equation
pouted bus.
e3811 note for Alabaster 1967.0
"Results
did not ditto so Ott were combined.
59
Appendix II-C/419
THERMAL TABLES-Continued
AccOmition
?
log time= al-b (temp.)
?
Dale Omits?
Lethal
Species
?
Stage/age?
Length?
Weight?
Sex?
Location?
Reference?
Extreme ?
?
(°C)?
1.050?
threshold+
Tamp
a
?Time?
a?
h?
Mb
?fil?
(%)
upper lower
Salvetinus full. Yearling
?
?
X =7.88 8?
Mixed
?
Codrington,?
Fry, Hart &?
Upper
naffs (brook
?
range 2-?
Ont. (hatch-?
Walker
trout)?
25 g?
cry
(1946)11
Uranus lonti-?
Juvenile?
Ontario,?
Fry and Gib-?
Upper
nalis (nanraycush
?
Canada?
son (1953)
42
hybrid)
Salvelinus?
1-2 yr. old?
? 27.7 gm ave. Mixed?
Hatcheries in Gibson and
?
Upper
namayoush?
(1 yr) 82.8?Ontario?
Fry (1954)11
(Lake trout)
?
gm M.
(2 yr)
Scardinius?
Adult ?10 cm?
?
Mixed?
Britain (field) Alabaster &
?
Upper
erythrophthala-?
Downing
mus (rudd)?
(1966)"
Semetilus afro-?
Adult?
2.0-3.9 ?
gm?
Mixed?
Don River,?
Hatt (1947)17?
Upper
maculates?
mode?
Thornhill,
(Creek chub)?
Ontario
Lower
Semotitus afro-?
Adult?
?
Toronto,
?
Hart (1952)11?
Upper
maculates?
Ontario
(Creek chub)?
Knoxville,
Tenn.
Sphaeroidis annu- Adult?
?
Northern Gull Heath (1967)11?
Upper
talus (Puffer)?
of Calif.
Coast
3
13.4325
11
14.6256
15
15.1846
20
15.0331
22
17.1967
24
17.8467
25
11.8467
10
13.2634
15
16.9596
20
19.4449
8?
1 wk
14.4820
15?
,
??
14.5123
20?
,,,,
11.3684
20
??
26.9999
5.
42.1859
10
31.0155
15
20.8055
20 ??
21.0214
25
16.8951
20
25
10 (Toronto only)
15 (Toronto only)
20.8055
20 (Toronto only)
19.1315
25
19.3186
30
22.8982
32.0
25.4649
-0.4556?
3?
-0.9991 26.0 23.5 ??
23.5
-0.4728?
6?
?
28.0 25.0
??
24.6
-0.4833?
9?
?
28.5 25.5
??
25.0
-0.4661?
7?
?
29.0 25.5 ??
25.3
-0.5367?
6?
?
29.0 26.5 ??
25.5
-0.5567 10?
?
30.0 25.5 ??
25.5
-0.5561?
1??
29.0 26.0 ??
25.5
-0.4381
?
6?
-0.9852 26.5 24.0 ?
23.5-24.0
-0.5540?
8?
-0.9652 28.0 24.5 ??
?
-0.6342?
9
?
-0.9744 28.0 24.5 ?
24.0-24.5
-0.5142?
4?
-0.9936 26?
23?
22.1
-0.4866?
5?
-0.9989 21?
24?
23.5
-0.5818
?
5?
-0.9951 21?
24?
23.5
-0.7692?
2< ?
-1.6021?
3?
-0.9408 26.0 25.0 ??
24.1
-1.0414?
3?
-0.8628 29.0 28.0 ??
27.3
-0.6226?
3?
-0.9969 31.0 30.0 ??
29.3
-0.5933?
1?
-0.9144 33.5 30.5 ??
30.3
-0.4499?
9?
-0.9911 35.0 31.0 ??
30.3
0.1
4.5
?
29?
28?21.5
-0.6226
?
3?
-0.9969 31?
30?
29
-0.5328?
6?
-0.9856 33?
30.5
??
30.5
-0.4111 18?
-0.9921 36?
32 ??
31.5
-0.5844 19?
-0.9961 31?
33
??
31.5
-0.6088?
3?
-0.9116 37.0 36.0
Sphaeroides mace-?" 13.8-15.9 cm 62.3-19.3 gm Mixed
?
New Jersey?
Hoff and West-?
Upper?
10?
11.3999 -0.2121?
3?
-0.9988 30.0 25.0-
??
21.5
fates (Northern?
(average)?
(average)? (40 N)?
man (1966)
50
?14?
35.5191 -1.0751
?
3?
-0.9449 32.0 21.0 ??
30.2
puffer)?
21?
21.5353 -0.5146?
3?
-0.9914 32.0 30.0 ??
31.2
28
? 23.7512 -0.6183
?
3?
-0.9239 33.5 31.1 ??
32.5
Lower
?
14??
-1.1104
?
0.6141?
4?
0.9760 10.0?
6.0 ??
1.1
21?
?
-3.5939?
0.7300
?
6?
0.9310 12.0?
8.0 ??
10.1
28
?
?
-7.4513 ' 0.8498
?
5?
0.9738 16.0 10.0
??
13.0
Thaleichthys?
Sexually?
161 mm M. 31 gm ave.?
Mixed?
Cowlitz River, Blahm &?
Upper
?
5 •?
river temp.
?
7.7440 -0.2140?
7?
-0.9142 29.0?
8.0 ??
10.5
pactrices?
Mature
?
Wash.?
McConnell
(Eulachon or
?
(1910)110
Columbia River?
unpublished
Smelt)?
data
Titania mossam- 4 months?
8.0-12.0 cm 10.0-17.0 gm ?
Transvaal?
Allanson &?
Upper?
22?
?
313.3130 -8.3878
?
4?
-0.8898 37.10 36.5
??
36.94 -
bin (Mourn-?
Africa?
Noble?
26?
14.0451 -0.2800?
5?
-0.2140 31.92 37.5
??
31.1
bique mouth-
?
(1914)71?
28
? 41.1610 -0.9950
?
4?
-0.3107 38.09 31.9 ?
?
31.81
breeder)
?
29
?
94.8243 -2.4125
?
5?
-0.1781 38.10 37.0
??
31.91-
30?
41.3233 -1.0018?
6?
-0.9124 38.50 37.6
??
31.51
32
? 34.0769 -0.8123?
4?
-0.9209 38.4 31.6
??
31.6
34??
123.1504 -3.1223
?
3?
-0.9938 38.4 38.2
?
. 31.25
36
?
68.6164 -1.7094
?
6?
-0.9053 38.71 37.9 ??
31.2
Tinca tinca?
Juvenile?
4.6-0.4 cm
?
Mixed?
England?
Alabaster &?
Upper?
15? 33.2000
?
1.0000?2. ?
(tench)
?
Downing11?
20? 29.6661
?
0.8333?
3
(1966)?
25? 21.1429
?
0.1143?
2
a
It is assumed in this table that the acclimation temperature reported is a true acclimation in the context of Brett
?
< Correlation coefficient (perfect Bt of all data points to the regression Ilne= 1.0).
(1952).71?
d =
Incipient lethal temperature of Fry, et al., (1946).11
Number of median resistance times used for calculating regression equation.
?
See previous note for Alabaster 1961.11
60
APPENDIX II-C
88
Alabaster, J. S. (1967), The survival of salmon
(Salmo solar
L.)
and sea trout
(S. trutta
L.) in fresh and saline water at high
temperatures.
Water Res.
1 (10):717-730.
69 Alabaster, J. S. and A. L. Downing (1966), A field and laboratory
.
investigation of the effect of heated effluents on fish.
Fish. MW.
Agr. Fish Food (Great Britain) Ser. I Sea Fish
6(4):1-42.
79
Alabaster, J. S. and R. L. Welcomme (1962), Effect of concentration
of dissolved oxygen on survival of trout and roach in lethal
temperatures.
Nature
194:107.
7 '
Allanson, B. R. and R. G. Noble (1964), The high temperature
tolerance of Tilapia mossambica (Peters).
Trans. Amer. Fish.
Soc.
93(4):323-332.
79
Allen, K. 0. and K. Strawn (1968), Heat tolerance of channel
catfish
Ictalurus punctatus,
in
Proceedings of the 21st annual conference
of the Southeastern Association . of Game and Fish Commissioners
(The
Association, Columbia, South Carolina), pp. 399-411.
73
Bishai, H. M. (1960), Upper lethal temperatures for larval sal-
monids. j.
Cons. Cons. Perma. Int. Explor. Mer
25(2):129-133.
74
Brett, J. R. (1952), Temperature tolerance of young Pacific sal-
•
mon, genus
Oncorhynchus. J.
Fish. Res. Board of Can., 9(6):
265-323.
7
6 Coutant, C. C. (1972),
Time-temperature relationships for thermal
resistances of aquatic organisms, principally fish
[ORNL-EIS 72-27]
Oak Ridge National Laboratory, Oak Ridge, Tennessee.
76
Coutant, C. C. (1970), Thermal resistance of adult coho salmon
(Oncorhynchus kisutch)
and jack chinook
(0. tshawylscha)
salmon
and adult steelhead trout
Salmo gairdneri
from the' Columbia
River. AEC Rept. No. BNWL-1580, Batelle Northwest, Rich-
land, Wash.
Craigie, D. E. (1963), An effect of water hardness in the thermal
resistance of the rainbow trout,
Salmo Gairdnerii, Can.
.7.
Zool.
41(5):825-830.
•
79
Doudoroff, P. (1942), The resistance and acclimatization of marine
fishes to temperature changes. I. Experiments With
Girella
nigriians
(Ayres).
Biol. Bull.
83(2):219-241.
•
79
Doudoroff, P. (1945), The resistance and acclimatization of marine
fishes to temperature changes. II. Experiments with
Fundulus
and
Atherinops. Biol. Bull.
88(2):194-206.
8
°
Edsall, T. A., D. V. Rottiers, and E. H. Brown (1970), Tempera-
ture tolerance of bloater
(Coregonus hoyi). J. Fish. Res. Board Can.
27(11):2017-2052.
81
Fry, F. E. J., J. R. Brett and G. H. Clawson (1942) Lethal limits
of temperature for young goldfish. Rev. Can. Biol. 1:50-56. -
•
"Fry,
F. E. J., and M. B. Gibson (1953), Lethal temperature ex-
periments with speckled trout x lake trout hybrids.
3. Hered.
44(2):56-57.
83
Fry, F. E. J., J. S. Hart and K. F. Walker (1946), Lethal tem-
peratures relations for a sample young speckled trout,
Salvelinus
fontinalis.
PM. Ont. Fish. Res. Lab. No. 66; Univ. of Toronto
Stud., Biol. Ser. No. 54, Univ. of Toronto press.
" Garside, E. T. and C. M. Jordan (1968), Upper lethal tempera-
tures at various levels of salinity in the euryhaline Cyprinodon-
tids
Fundulus heteroclitus
and
F. diaphanus
after isosomotic acclima-
tion.
3. Fish. Res. Board Can.
25(12):2717-2720.
" Gibson, E. S. and F. E. J. Fry (1954), The performance of the
lake trout,
Salvelinus namaycush,
at various levels of temperature
and oxygen pressure.
Can.
,
7.
Zool.
32(3):252-260.
66 J. R. (1971), Upper lethal temperature and thermal shock
tolerances of the opossum shrimp,
Neomysis awatschensis,
from
the Sacramento-San Joaquin estuary, California.
Calif. Fish
Game
57(0:17-27.
" Hart, J. S. (1947), Lethal temperature relations of certain fish of
the Toronto region.
Trans. Roy. Soc. Can.
Sec. 5(41):57-71.
" Hart, J. S. (1952),
Geographic variations of some physiological and
morphological characters in certain freshwater fish
[University of
Toronto biology series no. 60] (The University of Toronto
Press, Toronto), 79 p.
References Cited
1
"
Blahm, T. H. and R. J. McConnell,
unpublished data
(1970),
Mortality of adult eulachon
Thaleichthys pacificus
chinook slamon
and coho salmon subjected to sudden increases in water tem-
perature. (draft). Seattle Biological Laboratory, U.S. Bureau of
Commercial Fisheries, Seattle.
101 Blahm, T. H. and W. D. Parente,
unpublished data
(1970), Effects
of temperature on chum salmon, threespine stickelback and •
yellow perch in the Columbia river, Seattle Biological Labora-
tory, U.S. Bureau of Commercial Fisheries, Seattle.
Edsall, T. A. and P. A. Colby (1970), Temperature tolerance of
young-of-the-year cisco, Coregonus artedii. Trans. Amer. Fish.
Soc. 99(3):526-531.
193 McConnell, R. J. and T. H. Blahm,
unpublished data
(1970),
Resistance of juvenile sockeye salmon
0. nerka
to elevated water
temperatures. (draft) Seattle Biological Laboratory, U.S. Bureau
of Commercial Fisheries, Seattle.
104
Smith, W. E.
unpublished data
(1971), Culture reproduction and
temperature tolerance of
Pontoporeia affinis
in the laboratory.
(draft) National Water Quality Laboratory, Duluth, Minnesota.
106
G. R. and T. H. Blahm,
unpublished data
(1970), Mor-
tality of juvenile chinook salmon subjected to elevated water
temperatures. (draft Man.) Seattle Biological Laboratory. U.S.
Bureau of Commercial Fisheries, Seattle.
" Heath, W. G: (1967), Ecological significance of temperature tol-
erance in Gulf of California shore fishes.
.7.
Ariz. Acad. Sci.
4(3):I72-178.
" Hoff, J. G. and J. R. Westman (1966), The temperature tolerances
of three species of marine fishes.
J. Mar. Res.
24(2):131-140.
Lewis, R. M. (1965), The effect of minimum temperature on the
survival of larval Atlantic menhaden
Brevoortia tyrannus. Trans.
Amer. Fish. Soc.
94(4):409-412.
99
Lewis, R. M. and W. F. Hettler, Jr. (1968), Effect of temperature
and salinity on the survival of young Atlantic menhaden,
Bre-
voortia tyrannus. Trans. Amer. Fish. Soc.
97(4):344-349.
" McCauley, R. W. (1958), Thermal relations of geographic races of
Salvelinus. Can.
3.
Zool.
36(5):655-662.
94
McCauley, R. W. (1963), Lethal temperatures of the develop- .
mental stages of the sea lamprey,
Petromyzon marinus
L.
5.
Fish.
Res. Board Can.
20(2):483-490.
96
W. H., Jr., K. Strawn, and J. E. Dunn (1966), Heat resist-
ance experiments with the longear sunfish,
Lepomis miegalotis
(Rafinesque).
Arkansas Acad. Sci. Proc.
20:39-49.
9' Scott, D. P. (1964), Thermal resistance of pike
(Esox lucius L.)
muskellunge
(E. masquinongy)
Mitchill, and their F1 hybrids.
5.
Fish. Res. Board Can.
21(5) :1043-1049.
97
Simmons, H. B. (1971), Thermal resistance and acclimation at
various salinities in the sheepshead minnow
(Cyprinodon variegalus
Lacepede). Texas A&M Univ. Soc. No. TAMU-SG-71-205.
" Smith, W. E. (1970), Tolerance of
Mysis relicta
to thermal shock
and light.
Trans. Amer. Fish. Soc.
99(2):418-422.
99
Strawn, K. and J. E. Dunn (1967), Resistance of Texas salt- and
freshwater marsh fishes to heat death at various salinities,
Texas-T. Series, 1967:57-76.
Gi
APPENDIX C (ALL DATA ARE IN ° C)
FISH TEMPERATURE DATA
Species:
Alewife,
AZosa pseudoharengus
acclimation
I. Lethal threshold:
?
temperature
larvae?
juvenile
?
adult
Upper
5
15
5
10
20
5
15
23
5
20
23
5
*ultimate incipient?
32*
2
Lower
reference'
H. Growth:
Optimum and
[range]
larvae
juvenile?
adult
Ill. Reproduction:
optimum
range
month(s)
Migration
13*(3)
<10(1)-?
1.3
Spawning
16-28(1)
Apr-Aug(5)
1,5
17
11-27
1
Incubation
and hatch
*peak run
acclimation
IV.?
Preferred:
temperature
?
larvae
?
juvenile?
adult
24
23*
2
31
23*
2
18?
20
4
21?
22
*age unknown
'References on following page.
62
Alewife
References
1.
Edsall, T. A. 1970. The effects of temperature on the rate of development
and survival of alewife eggs and larvae. Trans. Amer. Fish. Soc. 99:376-
380.
2.
Carroll, E. W. and C. R. Norden. 1971. Temperature preference of the fresh-
water alewife, Alosa
pseudoharengus.
Abst. of paper presented at 33rd Midwest
Wildlife Conference.
3.
Tyus, H. M. 1974. Movements and spawning of anadromous alewives,
Alosa
pseudoharengus
(Wilson) at Lake Mattamuskeet, North Carolina. Trans. Amer.
Fish. Soc. 103:392-396.
4.
Meldrim, J. W., J. J. Gift, and B. • R. Petrosky. 1974. Supplementary data
on temperature preference and avoidance responses and shock experiments
with estuarine fishes and macroinvertebrates. Ichthyological Associates,
Inc., Middletown, Delaware. 56 p. mimeo.
5.
Graham, J. J. 1956. Observations on the alewife,
Pomolobus pseudoharengus
(Wilson), in fresh water. Univ. of Toronto, Biol. Ser. No. 62:43 p.
63
FISH TEMPERATURE DATA
Species:
Atlantic salmon,
Salmo salar
I. Lethal threshold:
acclimation
temperature larvae
?
juvenile?
adult reference'
Upper
5
22*
1
6
22
1
1 0
23*
1
20
23*
1
27.5
27.8**
Lower
*30 days after hatch
**ultimate upper incipient t
mp.
II. Growth:
larvae
juvenile
adult
Optimum and
10(9)
16-18(4)
4,9
[range]
Ill. Reproduction:
optimum
range
month(s)
Migration
adults 23 or less, smolt 10 or less
3
Spawning
4-6(3)
2-10(11)
Oct-Dec(7)
3,7,11
Incubation
3(3)-11(12)
3,12
and hatch
acclimation
IV.
?
Preferred:
temperature
larvae?
juvenile adult
4
14
2
Summer
17(5)
14-16(6) 5,6
14
10
References on following page.
Atlantic salmon
References
1. Bishai,, H. M.' 1960. Upper lethal temperatures for larval salmonids.
Jou. Du Conseil. 25:129-133.
2. Fisher, Kenneth C. and P. F. Elson. 1950. The selected temperature of
Atlantic salmon and speckled trout and the effect of temperature on the
response to an electrical stimulus. Physiol. Zoology. 23:27-34.
3. Dexter, R. 1967. Atlantic salmon culture. U.S. Bur. Sport Fish. Wildi.,
Mimeo.
4.
Markus, H. C. 1962. Hatchery reared Atlantic salmon smolts in ten months.
Prog. Fish. Cult. 24:127-130.
5.
Javoid, M. Y. and J. M. Anderson. 1967. Thermal acclimation and temperature
selection in Atlantic salmon,
Salmo salar,
and rainbow trout,
S. gairdneri.
J. Fish. Res. Bd. Canada.?
24(7):1515-1519.
6.
Ferguson, R. G. 1958. The preferred temperature of fish and their midsummer
distribution in temperate lakes and streams. J. Fish. Res. Bd. Canada.
15:607-624.
7.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology. Vol. 1.
Life History Data on
Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
8.
Garside, E. T. 1973. Ultimate upper lethal temperature of Atlantic salmon,
SaZmo saZar
L. Can. J. Zool. 51:898-900.
9.
Marr, D. H. A. 1966. Influence of temperature on the efficiency of growth
of salmonid embryos. Nature (London). 212:957-959.
10.
Legett, W. C. and G. Power. 1969. Differences between two populations of
landlocked Atlantic salmon
(Salmo saZar)
in Newfoundland. J. Fish. Res. Bd.
Canada. 16:1585-1596.
11.
Jones, J. W. 1959. The Salmon. Collins Press, London. 192 p.
12.
Spaas, J.
T. and M. J. Heuts. 1958. Contributions to the Comparative
Physiology and Genetics of the European Salmonidae. II. Physiologie et
Gênótique du Developpement Embryonnaire. Hydrobiologia. 12:1-26.
65
FISH TEMPERATURE DATA
reference'
Species:
Bigmouth buffalo,
Ictiobus cyprineilus
acclimation
I.
Lethal threshold: temperature larvae
?
juvenile
?
adult
Upper
Lower
II. Growth:
?
larvae
?
juvenile,
?
adult
Optimum and
[range)
III. Reproduction:
Migration
optimum
range
month(s)
Spawning
16-18(6)
14(1)-27(6)
?
Apr(4)-June(3)
1,3,4,6
2,5
Incubation
and hatch
141.5)-17(2,5)
acclimation
IV.
?
Preferred:
temperature
larvae?
juvenile
adult
11
-14*
7
*Ictiobus
sp. field
References on following page.
Bigmouth buffalo
References
1. Canfield, H. L. 1922. Cited in: Johnson, R. P. 1963. Studies on the
life history and ecology of the bigmouth buffalo,
Ictiobus cyprinelZus
(Valenciennes). J. Fish. Res. Bd. Canada. 20:1397-1429.
2.
Eddy, S. and J. C. Underhill. 1974. Northern Fishes. University of
Minnesota Press, Minneapolis. 414 pp.
3.
Walburg, C. H. and W. R. Nelson. 1966. Carp, river carpsucker, smallmouth
buffalo and bigmouth buffalo in Lewis and Clark Lake, Missouri River. U.S.
Bur. Sport Fish. Wildl., Washington, D.C. Research Report 69. 29 p.
4.
Harlan, J. R. and E. B. Speaker. 1956. Iowa Fish and Fishing. State
Conservation Commission. 377 p..
5. Walker, M. C. and P. T. Frank. 1952. The propagation of buffalo. Prog.
Fish. Cult.?
14:129-130.
6.
Swingle, H. S. 1957. Revised procedures for commercial production of
bigmouth buffalo fish in ponds in the southeast. Proc. 10th Ann. Conf.
S.E. Assoc. Game and Fish Comm. 1956. p. 162-165.
7. Gammon, J. R. 1973. The effects of thermal inputs on the population of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Research Center, Lafayette, Indiana. Tech. Rept. No. 32.
106 p.
67
1,5
6
*50% catch/effort
FISH TEMPERATURE DATA
Species:
Black crappie,
Pomoxis nigromaculatus
acclimation
I.
Lethal threshold: temperature
?
larvae?
juvenile
?
adult
Upper?
29
?
33*
Lower
*Ultimate incipient level
II.
Growth:?
larvae
?
juvenile?
adult
Optimum and?
22-25
[range]
?
(11-30)*
*Limits of zero growth
III.
Reproduction:
?
optimum
?
range?
month(s)
reference'
2
14(4)-20(3) Mar(4)-July(3)
Migration
Spawning
Incubation
and hatch
acclimation
IV. Preferred:
?
temperature larvae
?
juvenile
?
adult
Summer ?
18-20(5)?
24-34(1)
27-29*
References on following page.
68
Black crappie
References
1. Neill, W. H., J. J. Magnuson and G. G. Chipman. 1972. Behavioral thermo-
regulation by fishes - new experimental approach. Science. 176:1442-1443.
2.
Hokanson, K. E. F., and C. F. Kleiner. Effects of constant and diel
fluctuations. An temperature on growth and survival of black crappie.
Unpublished data, U. S. Environmental Protection Agency, Duluth, Minnesota.
3.
Breder, C. M., and D. E. Rosen. 1966. Modes of reproduction in fishes.
Nat. History Press. Garden City, New York. 941 p.
4.
Goodson, L. F. 1966. Crappie: In: Inland Fisheries Management, A.
Calhoun, ed., Calif. Dept. Fish and Game, p. 312-332.
5.
Faber, D. J. 1967. Limnetic larval fish in northern Wisconsin lakes.
J. Fish. Res. Bd. Canada. 24:927-937.
6.
Neill, W. H., and J. J, Magnuson. 1974. Distributional ecology and be-
havioral thermoregulation of fishes in relation to heated effluent from
a power plant at Lake Monona, Wisconsin. Trans. Amer. Fish. Soc., 103:
663-710.
69
FISH TEMPERATURE DATA
Species: ?
Bluegill,
Lepomis macrochirus
acclimation
• I. Lethal threshold: temperature larvae
?
juvenile?
adult
reference
Upper
15(2),
?
12(8)
27(8)
31(2)
2,8
20
32
2
25(2),
?
26(
.
8)
36(8)
33(2)
2,8
30
34
2
33
37
8
Lower
15(2),.12(8)
3?
(8)
3(2)
2,8
20
.
5
2
25(2),?
26(8)
10(8)
7(2)
2,8
30
11
2
33
15
8
II. Growth:
larvae
juvenile
adult
Optimum and
30(10)
24-27(3)
3,10
[range]
(22-34)(10)
[16(1)-30(4)1
III.
Reproduction:
?
optimum?
range
?
month(s)
Migration
Spawning
?
25(5)?
19(5)-32(6)?
AN-
Incubation
and hatch
?
22-24
?
22-34
acclimation
IV. Preferred:?
temperature larvae
?
juvenile adult
1,5,6
8
26 Aug(11)
32(9,11)
8 Nov
18
3 Feb
16
26 June
30 June
31
32
9,11
11
11
11
7
References on following page.
70
Bluegill
References
1. Emig,
J.
W. 1966. Bluegill sunfish.
?
In: Inland Fisheries Management.
A. Calhoun, ed., Calif. Dept. Fish and Game, p. 375-392.
2.
Hart, J. S. 1952. Geographical variations of some physiological and
morphological characters in certain freshwater fish. Univ. Toronto,
Biol. Ser. No. 60. 78 p.
3.
Anderson, R. O. 1959. The influence of season and temperature on growth
of
the
bluegill,
Lepomis macrochirus.
Ph.D. Thesis, Univ. Mich., Ann
Arbor. 133 p.
4. Maloney, John E. 1949. A study of the relationship of food consumption
of the bluegill,
Lepomis macrochirus
Rafinesque, to temperature. M.S.
Thesis, Univ. of Minn., Minneapolis. 43 p.
5.
Snow, H., A Ensign and John Klingbiel. 1966. The bluegill, its life
history, ecology and management. Wis. Cons. Dept., Madison. Publ. No.
230. 14 p.
6.
Clugston, J. P. 1966. Centrarchid spawning in the Florida Everglades.
Quart. J. Fla. Acad. Sci. 29:137-143.
7.
Cherry, D. S., K. L. Dickson, and J. Cairns, Jr. 1975. Temperatures
_selected and avoided by fish at various acclimation temperatures. J.
'Fish. Res. Bd. Canada. 32:485-491.
8.
Banner, A., and J. A. Van Arman. 1972. Thermal effects on eggs, larvae and
juveniles of bluegill sunfish. U. S. Environmental Protection Agency, Duluth,
Minnesota. Report No. EPA-R3-73-041.
9.
Ferguson, R. G. 1958. The preferred temperature of fish and their midsummer
distribution in temperate lakes and streams. J. Fish. Res. Bd. Canada.
15:607-624.
10.
Lemke, A. E. 1977. Optimum temperature
for
growth of it/Yen-He bluegills,
Lepomis macrochirus
Rafinesque. Prog. Fish Culturist, In press,
11.
Peterson, S. E., R. M. Schutsky, and S. E. Allison. 1974,
Temperature
pref-
. erence, avoidance and shock experiments with freshwater fishes and crayfishes.
Ichthyological Associates, Inc., Drumore, PA. Bulletin 10.
71
FISH TEMPERATURE DATA
Species:
Brook trout,
Salvelinus fontinalis
acclimation
I. Lethal threshold: temperature?
larvae
juvenile
?
adult
Upper
3
23
3
11
25
3
12
20*, 25**
2
15
25
3
20
*Newly hatched
25
3
Lower
25
**Swimup
25
3
II. Growth:
larvae
juvenile
adult
Optimum and
12-15(2)
16(1)
1,2
[range]
(7-18)(2)
(10-19)(1) 1,2
III.
Reproduction:?
optimum?
range
?
month(s)
Migration
Spawning
?
<9(.1)?
4 (6)-12(1)
?
6gEt -
Incubation
and hatch
?
6
?
?-13
acclimation
IV.
Preferred:
?
temperature larvae?
juvenile adult
6?
12
24
?
19
reference'
1.5,6
1
References on following page.
72
Brook trout
References
1.
Hokanson, K. E. F., J. H. McCormick, B. R. Jones, and J. H. Tucker. 1973.
Thermal requirements for maturation, spawning and embryo survival of the
brook trout,
Salvelinus fontinaZis
(Mitchill). J. Fish. Res. Bd. Canada.
30(7):975-984.
2.
McCormick, J. H., K. E. F. Hokanson, and B. R. Jones. 1972. Effects of
temperature on growth and survival of young brook trout,
Salvelinus
fontinalis.
J. Fish. Res. Bd. Canada. 29:1107-1112.
3.
Fry, F. E. J., J. S. Hart, and K. F. Walker. 1946. Lethal temperature
relations for a sample of young speckled trout,
Salvelinus fontinalis.
Univ. Toronto Studies, Biol. Ser. 54, Publ. Ontario Fish Res. Lab.
66:1-35.
4.
Cherry, D. S., K. L. Dickson, and J. Cairns, Jr. 1975. Temperatures
selected and avoided by fishes at various acclimation temperatures.
J. Fish. Res. Bd. Canada. 32:485-491.
5.
McAfee, W. R. 1966. Eastern brook trout. In: Inland Fisheries
Management, A. Calhoun, ed. Calif. Dept. Fish and Game. p. 242-264.
. Eddy, S., and J. C. Underhill. 1974. Northern Fishes. University of
Minnesota Press, Minneapolis. 414 p.
73
FISH TEMPERATURE DATA
Species:
Brown bullhead,
Ictalurus nebulosus
acclimation
I.
Lethal threshold: temperature
larvae?
juvenile?
adult
Upper
?
30?
35
Lower
II. Growth:?
larvae?juvenile?
adult
Optimum and
[range]
reference'
5
III. Reproduction:
optimum
range?
month(s)
Migration
Spawning
21(4)-?
?
Mar-Sept(3)
3,4
21(4)-27(3)
3,4
Incubation
and hatch
acclimation
IV.?
Preferred:
temperature
larvae
?
juvenile
?
adult
18 May(2)
21(2)?
29-31*(1)
1,2
26 July
31
2
23 Sept
27
2
10 Mar
26
2
*final preferendum
I
References on following page.
74
Brown bullhead
References
1.
Crawshaw, L. I. 1975. Attainment of the final thermal preferendum in
brown bullheads acclimated to different temperatures. Comp. Biochem.
Physiol.?
52:171-173.
2.
Meldrim, J. W., J. J. Gift, and B. R. Petrosky. 1974. Supplementary data
on temperature preference and avoidance responses and shock experiments
with estuarine fishes and macroinvertebrates. Ichthyological Associates,
Inc., Middletown, Delaware. 56 p. mimeo.
3.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
4
.
. Scott, W. B., and E. J. Crossman. 1973. Freshwater Fishes of Canada. J.
Fish. Res. Bd. Canada, Ottawa, Ontario. Bull. 184. 966 p.
5. Hart, J. S. 1952. Geographical variations of some physiological and
morphological
,
characters in certain freshwater fishes. Univ. Toronto
Biol. Ser. No. 60. 78 p.
75
FISH TEMPERATURE DATA
Species:
Brown trout,?
*ru**(7.
I. Lethal threshold:
acclimation
temperature?
larvae?
juvenile adult
reference'
Upper
20(2)
23(2)
26*(5)
2,5
23
25**
4
20
25**
4
15
25**
4
ID
24**
4
5
22**
4
*approx.?ultimate upper incipient 1 thal
Lower
**age unknown
II. Growth:
larvae
juvenile
adult
Optimum and
7-19*
4
[range]
*ages 0-IV
III. Reproduction:
Migration
optimum
range,
month(s)
6-7
Spawning
7-9(11)
1(7)-13(8)
Oct(9)-Jan(10
7,8,9,10,11
7-12(4)
3,4
Incubation
and hatch
5(4)-15(3)
acclimation
IV.?
Preferred:
temperature
larvae?
juvenile?
adult
12-18
6
References on following page.
76
Brown trout
References
1.
Stuart, T. A. 1953. Water currents through permeable gravels and their
significance to spawning salmonids. Nature. 172:407-408.
2. Bishai, H. M. 1960. Upper lethal temperatures for larval salmonids.
Jour. du Conseil. 25:129-133.
3.
Staley, J. 1966. Brown trout. In: Inland Fisheries Management,
,
A.
Calhoun, ed. Calif. Dept. of Fish and Game. p. 233-242.
4.
Frost, W. E., and M. E. Brown. 1967. The Trout, Collins Press, London.
286 p.
5.
Spaas, J. T. 1960. Contribution to the comparative physiology and genetics
of .
the European'salmonidae. III. Temperature eesistance at different ages.
Hydrobiologia. 15:78-88.
6.
Tait, J.
S. 1958. Cited in: Ferguson, R. G. 1958. The preferred
temperature of fish and their midsummer distribution in temperate lakes
and streams. J. Fish. Res. Bd. Canada. 15:607-624.
7. Verilidub, M. F. . 1963. Cited in; Brown, H. W. 1974. Handbook.Of the Effects
of Temperature on Some North American 'Fishes, American Elect. Power. Service
Corp.,
Canton, Ohio.'
8.
National Technical Advisory Committee. 1968. Water Quality Criteria.
Fed. Water Poll. Control Admin. U. S. Department of the Interior. 234 p.
9.
O'Donnell, D. J., and W. S. Churchill. 1954. Cited in: Carlander, K. D.
1969. Handbook of Freshwater Fishery Biology, Vol. 1. Life History Data
on Freshwater Fishes of
the United States and Canada, Exclusive
of the
Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
10.
Carl, G. C. 1938. A spawning run
of
brown trout in the Corvichan River
system. J. Fish. Res. Bd. Canada Progr. Rep. Pac. 36:12-13.
11.
Scott, W. B., and E. J. Crossman. 1973. Freshwater Fishes of Canada.
J. Fish. Res. Bd. Canada, Ottawa, Ontario: Bull. 184. 966 p.
77
reference'
3
3
10
9
2,4,5
1,7
1
acclimation
I. Lethal threshold: temperature
Upper
?
20
26
25-27
larvae?
juvenile
?
adult
31-34*
36*
40-41
FISH TEMPERATURE DATA
Species:
?
Carp,
Cyprinus carpio
Lower
*24 hr. TL50
II. Growth:
?
larvae
?
juvenile
?
adult
Optimum and
[range]
?
(16-30)(9)
III. Reproduction:
Migration
Spawning
Incubation
and hatch
IV. Preferred:
optimum
?
range
?
month(s)
14(4)-26(2)?Mar-Aug(5)
17-22(7)?
?-33(1)
Limit for 10 min.. exposure of early embryo
is 35°
acclimation
temperature larvae?
juvenile?
adult
25-35
?
31-32
Summer
?
33-35
10
?
17
19-23(2)
References on following page.
6
8
6
78
Carp
References
1.
Frank, M. L. 1973. Relative sensitivity of different stages of carp to
thermal shock. Thermal Ecology Symposium, May 3-5, 1973, Augusta, Georgia.
2.
Swee, U. B., and H. R. McCrimmon. 1966. Reproductive biology of the carp,
Cyprinus carpio L.,
in Lake St. Lawrence, Ontario. Trans. Amer. Fish.
Soc. 95:372-380.
3.
Black, E. C. 1953. Upper lethal temperatures of some British Columbia
freshwater fishes. J. Fish. Res. Bd. Canada. 10:196-210.
4.
Sigler, W. F. 1958. The ecology and use of carp in Utah. Utah State
Univ., Ag: Experiment Station. Bull. 405. 63 p.
5.
Carlander, K. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
6.
Pitt, T. K., E. T. Garside, and R. L. Hepburn. 1956. Temperature
selection of the carp
(Cyprinus carpio
Linn.). Can. J. Zool. 34:555-557.
7.
Burns, J. W. 1966. Carp. In: Inland Fisheries Management, A. Calhoun,
ed. Calif. Div. Game and Fish, p. 510-515.
8.
Gammon, J. R. 1973. The effect of thermal inputs on the population of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Res. Center, Lafayette, Indiana. Tech. Rept. No. 32.
9.
Tatarko, K. I. 1965. Cited in Brown, H. W. 1974. Handbook of the
Effects of. Temperature on Some North American Fishes. American Elect.
Power Service Corp., Canton, Ohio.
10.
Horoszewicz, L. 1973. Lethal and "disturbing" temperatures in some
fish species from lakes with normal and artifically elevated temperatures
J. Fish. Biol.
?
5:165-181.
7 9
FISH TEMPERATURE DATA
5062S:
?
Channel catfish,
Ictalurus punctatus
I. Lethal threshold:
reference'
Upper
acclimation
temperature
larvae?
juvenile
?
adult
15
30*
2
25(2)
?
26(1)
37(11?
34(2)*
1,2
29
31
3
30
37
1
34
38?
.
1
Lower
*SR-122
grams
15
0
2
20
3
2
25
6
2
II. Growth:
larvae
juvenile
?
adult
Optimum and
29-30(3)
28-30(8)
3,8
[range]
(27-31)(3)
.(26-34)(4)
3,4
III. Reproduction:
Migration -
optimum
range?
month(s)
Spawning
27(5)
21-29(5)?
Mar(10)-July(6) 5,6,10
24-28(5)
5
Incubation
and hatch
acclimation
IV.?
Preferred:
temperature
larvae?
juvenile
?
adult
Summer
30-32*
7
2 Jan(11)
11(11)
?
32**(9)
9,11
22
35
11
29
35
*field
11
**14-hr. photo eriod
References on following page.
80.
Channel catfish
References
1.
Allen, K. 0., and K. Strawn. 1968. Heat tolerance of channel catfish,
Ictalurus punctuatus.
Proc. 21st Ann. Conf. S.E. Assoc. Game and Fish
Comm., 1967, p. 399-411.
2.
Hart, J. S. 1952. Geographical variations of some physiological and
morphological characters in certain freshwater fish. Univ. Toronto, Toronto,
Ontario. Biological Series No. 60.
3. West, B. W. 1966. Growth, food conversion, food consumption, and survival
at various temperatures of the channel catfish,
Ictalurus punctatus
(Rafinesque).
M.S. Thesis, Univ.'Ark., Tuscon, Ark.
4.
Andrews, J.
W., and R. R. Stickney.. 1.972. Interaction of feeding rate and
environmental temperature of growth, food conversions, and body composition
of channel catfish. Trans. Amer. Fish. Soc. 101:94-97.
5.
Clemens, H. P., and K. F. Sneed. 1957. The spawning behavior of the channel
catfish,
Ictalurus punctatus.
U. S. Fish. Wildl. Serv., Special Sci. Rept.
Fish No. 219.
6.
Brown, L. 1942. Propagation of the spotted channel catfish,
IctaZurus
Zacustris punctatus.
Kan. Acad. Sci..Trans. 45:311-314.
7.
Gammon, J. R. 1973. The effect of thermal inputs on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Res. Center, Lafayette, Indiana. Tech. Rept. 32. 106 p.
8.
Andrews,
J.
W., L. H. ,Knight, and T. Murai. 1972. Temperature requirements
for high density rearing of channel catfish from fingerling to market size.
Prog. Fish. Cult. 34:240-241.
9.
Kilambi, R. V., J. Noble, and C. E. Hoffman. 1970. Influence of temperature
and food conversion efficiency of the channel catfish. Proc. 24th Ann. Conf.
S.E. Assoc. Game and Fish Comm., 1969, p. 519-531.
10.
Stevens, R. E. 1959. The white and channel catfishes of the Santee-Cooper
Reservoir and Tailrace Sanctuary. Proc. 13th Ann. Conf. S.E. Assoc. Game
and Fish. Comm., 1959, p. 203-219.
11.
Peterson, S. E., R. M. Schutsky, and S. E. Allison. 1974. Temperature
preference, avoidance, and shock experiments with freshwater fishes and
crayfishes. Ichthyological Associates, Inc., Drumore, Pennsylvania.
Bull. 10.
81
FISH TEMPERATURE DATA
Species:
?
Coho salmon,
Oncorhynchus kisutch
reference'
acclimation
I. Lethal threshold:?
temperature
larvae
?
juvenile
adult
Upper
10
5
2324(1)
21*(3)
1
1,3
15
24
1
20
25
1
23
25
1
*Accl.?
temp.
nknown
•?
Lower
5
0.2
1
10
2
1
15
3
1
20
5
1
23
6
1
II. Growth:
larvae
juvenile
adult
Optimum and
15*
2
[range]
(5-17)**
III. Reproduction:
optimum
*unlimited food
**depending upon season
range
?
month(s)
Migration
7-16
Spawning
7-13
Fall
3
8(2)
?-11(7)
2,7
Incubation
and hatch
acclimation
IV.
?
Preferred:
temperature
larvae?
juvenile
adult
Winter
13
4
References on following page.
82
Coho salmon
References
1.
Brett, J. R. 1952. Temperature tolerance in young Pacific salmon, genus
Oncorhynchus.
J.
.Fish. Res. Bd. Canada. 9:265-323.
2.
Great. Lakes Research Laboratory. 1973. Growth of lake trout in the
laboratory. Progress in Sport Fishery Research. 1971. USDI, Fish and
Wildlife.
Service, Bureau of Sport Fisheries and Wildlife. p. 100 and 107.
3.
U. S. Environmental Protection Agency. 1971. Columbia River thermal
effects study, Vol. 1. Biological Effects Studies. 95 p.
4.
Edsall, T. 1970. U. S. Dept. of Int., Great.Lakes Fishery Laboratory,
Ann Arbor, Michigan. Personal Communication.
5.
Burrows, R. E. 1963. Water temperature requirements for maximum
productivity of salmon. Proc. 12th Pacific N. W. Symposium on Water
Poll. Res., Nov. 7, 1963, Corvallis, Oregon. p. 29-38.
6.
Averett, R. C. 1968. Influence of temperature on energy and material
utilization by juvenile coho salmon. Ph.D. Thesis, Oregon State Univ.,
Corvallis, Oregon.
7.
Shapovalov, L. and A. C. Taft. 1954. Cited in: Schuytema, G. 1969.
Literature review, effects of temperature on Pacific salmon, Appendix A.
In: Thermal Pollution: Status of the Art, Parker, F. L. and R. A. Krenkel,
ed. Vanderbilt Univ., Nashville, Tennessee. Rept. No. 3. 31.7 p.
83
FISH TEMPERATURE DATA
Species: ?
Emerald shiner,
Notropis atherinoides
acclimation
?
I. Lethal threshold:
temperature
?
larvae
juvenile
?
adult
reference
I
Upper
15
29
1
20
31
1
25
31
1
Lower
15
2
1
20
5
1
II. Growth:
Optimum and
larvae
juvenile,
adult
29
2
[range]
(24-31)
2
III. Reproduction:
Migration
Spawning
Incubation
and hatch
optimum
range
month(s)
20(3)-28(5)
May-Auq(1,4)
1,3,4,5
acclimation
IV. Preferred:
?
temperature larvae
?
juvenile
?
adult
Summer?
25*
*unknown age
I
References on following page.
3
84
Emerald shiner
References
1.
Carlander, R. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
2. McCormick, J. H., and C. F. Kleiner. 1976. Growth and survival of young-
of-the-year emerald shiners
(Notropis atherinoides)
at different
temperatures. J. Fish. Res. Bd. Canada. 33:839-842.
3. Campbell, J. S., and H. R. MacCrimmon. 1970. Biology of the emerald shiner
Notropis atherinoides
Rafinesque in Lake Simcoe, Canada. J. Fish. Biol.
2:259-273.
4.
Flittner, G. A. 1964. Morphometry and life history of the emerald shiner
Notropis
atherinDis .
Rafinesque. Ph.D. Thesis, Univ. of Mich., Ann Arbor,
Michigan.
5.
Gray, J. W. 1942. Studies on
Notropis atherinoides atherinoides
Rafinesque
in the Bass Islands Region of Lake Erie. M.S. Thesis, Ohio State Univ.,
Columbus, Ohio.
85
FISH TEMPERATURE DATA
Species:
?
Fathead minnow,
Pimephales promelas
acclimation
I.
Lethal threshold: temperature larvae
?
juvenile
?
adult
Upper
Lower
II. Growth:?larvae
?
juvenile?
adult
Optimum and
?
23.5-30
[range]
reference'
1
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
23.5(1)
18(2)-30(1)
May-Aug(2) 1,2
Incubation
and hatch
23-2.8
23.5-30
1
acclimation
IV.
Preferred:?
temperature larvae?
juvenile adult
I
References on following page.
86
Fathead minnow
References
1.
Brungs, W. A. 1971. Chronic effects of constant elevated temperature
on the fahead minnow
(Pimephales promeZas
.
Rafinesque). Trans. Am. Fish.
Soc. 100:659-664.
2.
Carlson, D. R. 1967. Fathead minnow,
Pimephales promeZas
Rafinesque, in
the Des Moines River, Boon County, Iowa, and the Skunk River Drainage,
Hamilton and Story Counties, Iowa. Iowa State J. Science. 41:363-374.
87
reference
FISH TEMPERATURE DATA
Species:
Freshwater drum,
Aplodinotus grunniens
acclimation
I. Lethal threshold: temperature larvae
?
juvenile
?
adult
Upper
Lower
II. Growth:
?
larvae
?
juvenile
?
adult
Optimum and
[range]
III. Reproduction:
optimum
range
?
month(s)
Migration
Spawning
18-24(4)?
May(1)-Auq(3)
1,3,4
Incubation
and hatch
22(2)-26(1)
1,2
acclimation
IV.?
Preferred:
temperature
larvae?
juvenile?
adult
29-31*
5
*Field
' References on following page.
88
Freshwater drum
References
1.
Wrenn, B. B. 1969. Life history aspects of smailmouth buffalo and
freshwater drum in Wheeler Reservoir, Alabama. Proc. 22nd Ann.
Conf. S.E. Assoc. Game and Fish Comm., 1967. p. 479-495.
2.
Davis, C. C. 1959. A planktonic fish egg from freshwater. Limn. Ocean.
4:352-355.
3.
Edsall, T. A. 1967. Biology of the freshwater drum in Western Lake Erie.
Ohio J. Sci. 67:321-340.
4.
Swedberg,
D.
V., and C. H. Walburg. 1970. Spawning and early life history
of the freshwater drum in Lewis and Clark Lake, Missouri River. Trans; Am.
Fish. Soc. 99:560-571.
5.
Gammon, J. R. 1973. The effect of,thermal inputs on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Research Center, Lafayette, Indiana. Tech. Rept. 32. 106 p.
89
FISH TEMPERATURE DATA
Species:
Lake Herring (cisco),
Coregonus artedii
acclimation
1. Lethal threshold: tem erature larvae
?
juvenile?
adult?
reference'
20(2)?
20(3)?
204)?
2,3,4
Upper?
22(3)?
<2
(
4 5
*
)?
3,5
26
?
3
20
? 26?
3
25
?
26?
3
*accl. temp. unknown
Lower
?
2?
.0.3?
3
5?
0.5
?
3
10?
3
?
3
20?
5?
3
25
?
10
?
3
II. Growth:?
larvae?
juvenile,
?
adult
Optimum and
?
16
[range]?
(13-1R)
III.
Reproduction:
?
optimum
?
range
?
month(s)
>13
2
2
To spawning grounds at
?
5
3(6,7)?
1-5(8)?
Nov-Dec(6)
Migration
Spawning
Incubation
and hatch
2-8(1)?
Nov(6)-May(8)
7
6,7,8
1,6,8
acclimation
IV. Preferred:?
temperature larvae
?
juvenile adult
13
6(
1 )
6
References on following page.
40
Lake herring (cisco)
References
1.
Colby, P. J., and L. T. Brooke. 1970. Survival and development of the
herring
(Coregonus artedii)
eggs at various incubation temperatures.
In: Biology of Coregonids, C. C. Lindsay and C. S. Woods, ed. Univ.
Manitoba, Winnipeg, Manitoba, Canada. pp. 417-428.
2.
McCormick, J. H., B. R. Jones, and R. F. Syrett. 1971. Temperature
requirements for growth and survival of larval ciscos
(Coregonus artedii).
J. Fish. Res. Bd. Canada.
•
28:924-927.
3. Edsall, T. A., and P. J. Colby. 1970. Temperature tolerance of young-of-
the-year cisco,
Coregonus artedii.
Trans. Amer. Fish. Soc. 99:526-531.
4. Frey, D. G. 1955. Distributional ecology
of
the cisco
(Coregonus artedii).
'Investigations of Indiana Lakes and Streams. 4:177-228.
5.
Colby, P. J., and L. T. Brooke. 1969. Cisco
(Coregonus artedii)
mortalities
in a Southern Michigan lake, July 1968. Limn. Ocean. 14:958-960.
6.
Dryer, W. R., and J. Beil. 1964. Life history of lake herring in Lake
Superior. U.S. Fish. Bull. 63:493-530.
7.
Cahn, A. R. 1927. An ecological study of southern Wisconsin fishes, the
brook siversides
(Labidesthes sicculus)
and . the cisco
(Leucichthys artedii,
LeSueur).
?
Ill. Biol. Monogr.. 11:1-151.
8.
Carlander, K. D. 1969. Handbook
of
Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes
of
the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
91
FISH TEMPERATURE DATA
Species:
Lake trout,
Sedvelinus namaycush
acclimation
I.
Lethal threshold: temperature larvae
?
juvenile
?
adult
Upper
Lower
II. Growth:?
larvae?
juvenile
?
adult
Optimum and
[range]
reference'
III. Reproduction:
Migration
optimum
range
month(s)
Spawning
3-14(3)
Aug-Dec(2)
2,3
8(
1)
0.3-10(3)
1,3
Incubation
and hatch
acclimation
IV.?
Preferred:
temperature
larvae
?
juvenile adult
12*
4
8-1S**
5
*yearling
**age unknown
References on following page.
9.2
Lake trout
References
1. Edsall, T. A., and W. E. Berlin. 1973. In: Progress in Fishery Research
1973, Eschmeyer, P. H., and J. Kutkuhn, eds. U. S. Fish and Wildlife
Service, Great Lakes Fishery Laboratory. Ann Arbor, Michigan.
2.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
3.
Royce, W. F. 1951. Breeding habits of lake trout in New York. Fishery
Bull. 52:59-76.
4.
McCauley, R. W., and J. S. Tait. 1970. Preferred temperature of yearling
lake trout,
Salvelinus namaycush.
J. Fish. Res. Bd. Canada. 27:1729-1733.
5.
Ferguson, R. G. 1958. The preferred temperature of fish and their midsummer
distribution in temperate lakes and streams. J. Fish. Res. Bd. Canada.
15:607-624.
93
FISH TEMPERATURE DATA
Species:
Lake whitefish,
Coregonus clupeaformis
acclimation
I.
Lethal threshold:
temperature larvae?
juvenile
?
adult •
Upper
Lower
II.
Growth:?
larvae?
juvenile?
adult
Optimum and
[range]
reference'
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
0.5-10
Sept-Dec
2
Incubation
and hatch
3-8
1
acclimation
IV. Preferred:
temperature
larvae
?
juvenile
adult
13*
3
*2 year old
References on following page.
44
Lake whitefish
References
1.
Brooke, L. T. 1975. Effect of different constant incubation temperatures
on egg survival and embryonic development in lake whitefish
(Coregonus
clupeaformis).
Trans. Amer. Fish. Soc. 3:555-559.
2.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p.
3.
Ferguson, R. G. 1958. The preferred temperature of fish and their mid-
summer distribution in temperate lakes and streams. J. Fish. Res. Bd.
Canada. 15:607-624.
95
FISH TEMPERATURE DATA
Species:?
Largemouth bass,
Micropterus saimoides
I. Lethal threshold:
reference'
acclimation
temperature
larvae?
juvenile adult
Upper
20
33
1
25
35
1
30
36
1
Lower
20
5
25
•?
7
hes
V
S/
12
1
II. Growth:
larvae
juvenile
adult
Optimum and
27(2)
'30(8)
2,8
[range]
(20-30)(2)
{23-31)(8)
2,8
29(10)
22(11)
10,11
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
May(4)
21(4)
16-27(4)
Apr-June(3)Nov-May
3,4
and hatch
20(5)
13(6)-26(9)
5,6,9
acclimation
IV.
?
Preferred:
temperature
larvae?
juvenile
adult
30-32*
7
27-28**
7
?
?
*Lab., small
**Field, larger
References on following page.
96
Largemouth bass
References
1.
Hart, J. S. 1952. Geographic variations of some physiological and.
morphological characters in certain freshwater fish. Univ. Toronto,
Toronto, Ontario. Biological Series No. 60.
2.
Strawn, Kirk. 1961. Growth of largemouth bass fry at various temperatures
Trans. Amer. Fish. Soc. 90:334-335.
3.
Kramer, R. H., and L. L. Smith, Jr. 1962. Formation of year class in
largemouth bass. Trans. Amer. Fish. Soc. 91:29-41.
4.
Clugston,
J.
P. 1966. Centrarchid spawning in the Florida Everglades:
Quart. J. Fla. Acad. Sci. 29:137-143.
5.
Badenhuizen, T. 1969. Effect of incubation temperature on mortality of
embryos of largemouth bass,
Micropterus saZmoides
Lacepede. M.S. Thesis,
Cornell University, Ithaca, New York.
6. Kelley, J. W. 1968. Effects of incubation temperature on survival of
largemouth bass eggs. Prog. Fish. Cult. 30:159-163..
7.
Ferguson, R. G. 1958. The preferred temperature of fish and their •
midsummer distribution
in
temperate lakes and streams. J. Fish. Res.
Bd. Canada. 15:607-624.
8.
Lee, R. A. 1969. Bioenergetics of feeding and growth of largemouth bass
in aquaria and ponds. M.S. Thesis, Oregon State University, Corvallis,
Oregon.
9.
Carr, M. H. 1942. The breeding habits, embryology and larval development
of the largemouth black bass in Florida. Proc. New Eng: Zool. Club.
20:43-77.
10.
Johnson, M. G., and W. H. Charlton. 1960. Some effects of temperature on
metabolism and activity of largemouth bass
Micropterus saZmoides
Lacepede.
Prog. Fish. Cult. 22:155-163.
11.
Markus, H. C. 1932. Extent to which temperature changes influence food
consumption in largemouth bass
(Hero floridans).
Trans. Am. Fish. Soc.
62:202-210.
97
FISH TEMPERATURE DATA
Species:
?
Northern pike,
Esox Lucius
temperature
?
larvae
?
juvenile
?
adult reference'
Upper
18
25 28*
2
25
32
1
27
33
1
30
33**
1
*At hatch and free swimming, respe
ti
vely
Lower
**Ultimate incipient level
18
3*
II. Growth:
Optimum and
[range]
larvae
*At hatch and free swimming
juvenile
?
adult
21
26
(08-26)
2
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
4(4)-18(3)
Feb-June(5)
3,4,5
Incubation
and hatch
12
7-19
2
acclimation
IV.
?
Preferred:
temperature
larvae
?
juvenile
adult
24,26*
6
*Grass pickerel and musky,
respectively
References on following page.
Northern Pike
References
1. Scott, D. P. 1964. Thermal resistance of pike
(Esox Zucius L.),
muskellunge
(E. masquinongy,
Mitchell), and their F 1
hybrid. J. Fish.
Res. Bd. Canada. 21:1043-1049.
2. Hokanson, K. E. F'.,
J.
H. McCormick, and B. R. Jones. 1973. Temperature
requirements for embryos and larvae of the northern pike,
Esox Zucius
(Linnaeus). Trans. Amer. Fish. Soc. 102:89-100.
3.
Fabricus, E., and K. J. Gustafson. 1958. Some new observations on the
spawning behavior of the pike, Esox Zucius
L. Rep. Inst. Freshwater
Res., Drottningholm, Sweden. 39:23-54.
4.
Threinen, C. W., C. Wistrom, B. Apelgrn, and H. Snow. 1966. .The northern
pike, its life history, ecology, and management. Wis. Con. Dept., Madison,
Publ. No. 235. 16 p.
5.
Toner, E. D., and G. H. Lawler. 1969. Synopsis of biological data on the
pike
Esox Zucius
(Linnaeus 1758). Food and Ag. Org. Fisheries synopsis
No. 30, Rev. 1. Rome. 37 p.
6.
Ferguson, R. G. 1958. The preferred temperature of fish and their midsummer
distribution in temperate lakes and streams. J. Fish. Res. Bd. Canada.
15:607-624.
99
larvae
?
juvenile
?
adult
30
15-?
optimum
range
20-29
month(s)
May-Aug
FISH TEMPERATURE DATA
Species: ?
Pumpkinseed,
Lepomis gibbosus
acclimation
I. Lethal threshold: temperature
Upper
larvae?
juvenile?
adult
Lower
II. Growth:
Optimum and
[range]
III.
Reproduction:
Migration -
Spawning
Incubation
and hatch
reference'
acclimation
IV. Preferred:?
temperature
?
larvae
juvenile?
adult
19 May
21
2
24 June
31
2
26 Sept
33
2
8 Nov
10
2
References on following page.
100
Pumpkinseed
References
1.
Pessah, E., and P. M. Powles. 1974. Effect of constant temperature on
growth rates of pumpkinseed sunfish
(Lepomis gibbosus).
J.
Fish. Res.
Bd. Canada. 31:1678-1682.
2. Peterson, S. E., R. M. Schtusky, and S. E. Allison. 1974. Temperature
preference, avoidance and shock experiments with freshwater fishes and
crayfishes. Ichthyological Associates, Inc., Drumore, Pennsylvania.
Bulletin 10.
3. Breder, C. M., Jr. 1936. The reproductive habits of the North American
sunfishes (family centrarchidae). Zoologica. 21:1-48.
FISH TEMPERATURE DATA
Species:
?
Rainbow smelt,
Osmerus mordax
acclimation
I.
Lethal threshold: temperature larvae
?
juvenile?
adult
Upper
Lower
II.
Growth:
?
larvae
?
juvenile
?
adult
Optimum and
[range]
reference'
III. Reproduction:
Migration
Spawning
Incubation
and hatch
optimum
range
month(s)
4-5
1
0.6-15
April
5-15
acclimation
IV.
Preferred:?
temperature larvae
?
juvenile adult
6-14
I
References on following page.
4
102
Rainbow smelt
References
1.
McKenzie, R. A. 1964. Smelt life history and fishery in the Miramichi
River, New Brunswick. J. Fish. Res. Bd. Canada, Ottawa, Ontario. Bull.
144. 77 p.
2. Hale, J. G. 1960. Some aspects of the life history of the smelt
(Osmerus mordax)
in Western Lake Superior. Minn. Fish & Game Invest.
Fish Ser. 2:25-41.
3.
Carlander, K.?
1969. Handbook
of
Freshwater Fishery Biology, Vol. 1.
Life History on Freshwater Fishes of the United States and Canada, Exclusive
of the Perciformes. Iowa State. Univ. Press, Ames, Iowa. 752.p.
4.
Wells, L. 1968. Seasonal depth distribution of fish in southeastern Lake
Michigan. Fishery Bull. 67:1-15.
103
FISH TEMPERATURE DATA
Species:
?
Rainbow trout,
SaZmo gairdneri
acclimation
I. Lethal threshold: temperature larvae?
juvenile
?
adult
Upper?
18?
27
19?
21
Lower
reference'
1
II. Growth:
larvae
juvenile
adult
Optimum and
17-19
5
[range]
[3(8)
-20 (1 1) ]
8,11
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
9(10)
5-..13(6)
Nov-Feb(7)
6,7,10
In cu
bation
Feb-June
7)
and hatch
5-7(9)_
5-13(4)
4,9
acclimation
IV.
?
Preferred:
temperature
larvae?
juvenile
adult
Not given
14
3
13-20
?
13-19
11
18&24
18&22. reap.
12
References on following page.
104
Rainbow trout
References
1.
Alabaster, J. S., and R. L. Welcomme. 1962. Effect of concentration of
dissolved oxygen on survival of trout and roach in lethal temperatures.
Nature (London). 194(4823):107.
2. Coutant, C. C. 1970. Thermal resistance of adult coho
(Oncorhynchus kisutch)
and jack chinook
(O.
tshawytscha)
salmon, and the adult steelhead trout
(Salm° gairdnerii)
from the Columbia River. Atomic Energy Commission, Battelle
Northwest Laboratory, Richland, Washington, publication No. 1508, 24 p.
3.
Ferguson, R. G. 1958. The preferred temperature of fish and their midsummer
distribution in temperate lakes and streams. J. Fish. Res. Bd. Canada.
15:607-624.
4.
McAfee, W. R. 1966. Rainbow trout. In: Inland Fisheries Management, A.
Calhoun, ed. Calif. Dept. Fish and Game. pp. 192-215.
5.
Hokanson, K. E. F., C. F. Kleiner, and T. W. Thorslund. 1976. Effects of
constant temperature and diel fluctuation on specific growth, mortality,
and yield of juvenile rainbow trout,
SaZmo gairdneri
(Richardson). MS
submitted to J. Fish. Res. Bd. Canada.
6.
Rayner, H. J. 1942. The spawning migration of rainbow trout at Skaneateles
Lake, New York. Trans. Amer. Fish. Soc. 71:180-83.
7.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames, Iowa. 752 p. •
8.
Vojno, T. 1972. The effect of starvation and various doses of fodder on
the changes of body weight and chemical composition and the survival rate -
in rainbow trout fry
(Salm° gairdneri,
Richardson) during the winter.
Roczniki Nauk Rolniczych Series H - Fisheries 94, 125. In: Coutant, C. C.,
and H. A. Pfuderer. 1973. Thermal effects. J. Water Poll. Fed. 46:1476-1540.
9. Timoshina, L. A. 1972. Embryonic development of the rainbow trout
(Salmo
gairdneri irideus,
Gibb.) at different temperatures. Icthyol. (USSR).
12:425.
10.
Johnson, Charles E. 1971. Factors affecting fish spawning. Wisconsin
Cons. Bull. July-Aug.
11.
Mantelman,
?
I. 1958. Cited in: Brown, H. W. 1974. Handbook of the
Effects of Temperature on Some North American Fishes. American Elect.
Power Service Corp., Canton, Ohio.
12.
Cherry, D. S., K. L. Dickson, and J. Cairns, Jr. 1975. Temperatures
selected and avoided by fish at various acclimation temperatures. J.
Fish. Res. Bd. Canada. 32:485-491.
105
FISH TEMPERATURE DATA
Species:
?
Sauger,
Stizostedion canadense
I. Lethal threshold:
acclimation
temperature?
larvae
juvenile
?
adult
reference'
Upper
10
27
4
12
27
4
18
29
4
22
30
4
26
30
4
Lower
II. Growth:
Optimum and
[range]
larvae
juvenile
adult
22
(16-26)
4
III. Reproduction:
optimum
range
?
month(s)
Migration
Spawning
9-15(4)*
6(1)-15(4)?
Apr(1)-June(3)
1,3,4
Incubation
12-15
4
and hatch
9-18
*for fertilization
acclimation
IV. Preferred:
temperature
larvae
?
juvenile?
adult
19*
2
27-29
5
*field
References on following page.
106
Sauger
References
1. Nelson, W. R. 1968. .Reproduction and early life history of sauger,
Stizostedion canadense,
in Lewis and Clark Lake. Trans. Amer. Fish.
Soc. 97:159-166.
2. Ferguson, R. G. 1958. The preferred temperature of fish and their
midsummer distribution in temperate lakes and streams. J. Fish. Res.
Bd. Canada. 15:607-624.
3.
Carufel, Louis H. 1963. Life history of saugers in Garrison Reservoir.
J. Wildl. Manag. 27(3):450-456.
4.
Smith, L. L., Jr., and
W. M. Koenst. 1975. Temperature effects on eggs and
fry of percoid fishes. U. S. Environmental Protection Agency, Duluth,
Minnesota. Report EPA-660/3-75-017. 91 p.
?
•
5.
Gammon,
J. R. 1973. The effect of thermal input on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Res. Center, Lafayette, Indiana. Tech. Rept. 32. 106 p.
FISH TEMPERATURE DATA
Species:
Smallmouth bass,
Micropterus dolomieui
acclimation
I. Lethal threshold: temperature larvae
?
juvenile?
adult
Upper?
?
38*(8)?35(3)
*acclimation not given
4(R)*
?
2(3)
3,8
4
3
73
10
3
*acclimation temperature not giv
juvenile
?
adult
26(3)
2,3
Lower
?
15(3)
18
22
26
II. Growth:?
larvae
Optimum and
?
28-29(2)
[range]
III.
Reproduction:
?
optimum
Migration
Spawning
?
17-18(5)
Incubation
and hatch
acclimation
IV.
Preferred:
?
temperature
Summer
Winter
18&30
range?
month(s)
13-23(9)?
May-June(7)
5,7,9
13-22
10
larvae?
juvenile
?
adult
21-27
6
>8*(1)-28(4)
1,4
23&31 resp.
11
*juvenile and adult
I
References on following page.
108
Smallmouth bass
References
1.
Munther, G. L. 1970. Movement and distribution of smallmouth bass in
the Middle Snake River. Trans. Amer. Fish. Soc. 99:44-53.
2.
Peek, F. W. 1965. Growth studies of laboratory and wild population
samples of smallmouth bass,
Micropterus dolomieui
Lacepede, with
applications to mass marking of fishes. M.S. Thesis. Univ. Ark.,
Fayetteville. 115 p.
3
?
Horning, W. B., and R. E. Pearson. 1973. Growth temperature requirements
and lower lethal temperatures for juvenile smallmouth bass
(Micropterus
dolomieui).
J.
Fish. Res. Bd. Canada. 30:1226-1230.
4. Ferguson, R. G. 1958. The preferred temperature of fish and their
midsummer distribution in temperate
.
lakes and streams. J. Fish. Res.
Bd. Canada. 15:607-624.
Breder, C. M., and D. E. Rosen. 1966. Modes of reproduction in fishes.
Natural History Press, Garden City, New York. 941 p.
6... Emig, J. W. 1966. Smallmouth bass. In: Inland Fisheries Management,
A. Calhoun, ed. Calif. Dept. Fish and Game. pp. 354-366.
7.
Surber, E. W. 1943. Observations on the natural and artificial propaga-
tion of the smallmouth black bass,
Micropterus dolomieui.
Trans. Amer.
Fish. Soc. 72:233-245.
8.
Larimore, R.
W.,
and M. J. Duever. 1968. Effects of temperature acclimation
on the swimming ability of smallmouth bass fry. Trans. Amer. Fish. Soc.
97:175-184.
-9. Tester, A. L. 1931. Cited in: Wallace, C. R. 1973. Effects of tempera-
ture on developing meristic structures of smallmouth bass,
Micropterus
doZomieui
Lacepede. Trans. Amer. Fish. Soc. 102:142-144.
10.
Webster, D. A. 1945. Relation of temperature to survival and incubation
of the eggs of smallmouth bass
(Micropterus dolomieui).
Trans. Amer. Fish.
Soc. 75:43-47.
11.
Cherry, D. S., K. L. Dickson, and J. Cairns, Jr. 1975. Temperatures
selected and avoided by fish at various acclimation temperatures. J. Fish.
Res. Bd. Canada. 32:485-491.
109
FISH TEMPERATURE DATA
Species:
?
Smallmouth buffalo,
Ictiobus bubalus
acclimation
I. Lethal threshold: temperature larvae?
juvenile
?
adult
Upper.
Lower .
II.
Growth:?
larvae
?
juvenile
?
adult
Optimum and
[range]
III.
Reproduction:
?
optimum
?
range?
Month(s)
Migration
Spawning
Incubation
and hatch
17(1)-24(5)
14(1)-28(5) Mar(3)-Sept(5)
14(1)-21(2)
acclimation
IV. Preferred:
?
temperature larvae
?
juvenile adult
31-34*
*Ictiobus
sp. fiel
References on following page.
reference'
1,3,5
1,2
110
Smallmouth buffalo
References
1.
Wrenn, W. B. 1969. Life history aspects of smallmouth buffalo and
freshwater drum in Wheeler Reservoir, Alabama. Proc. 22nd Ann. Conf.
S.E. Assoc. Game & Fish Comm., 1968. pp. 479-495.
2.
Walburg, C. H., and W. R. Nelson. 1966. Carp, river carpsucker, small-
mouth buffalo and bigmouth buffalo in Lewis and Clark Lake, Missouri River.
U. S. Bur. Sport Fish. Wildl., Washington, D. C. Res. Rep. 69. 29 p.
3.
Walker, M. C., and P. T. Frank. 1952. The propagation of buffalo. Prog.
Fish. Cult.?
14:129-130.
4.
Gammon, J. R. 1973. The effect of thermal input on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Research Center, Lafayette, Indiana: Tech. Rept. 32. 106 p.
5.
Jester, D. B.- 1973. Life history, ecology, and management of the small-
mouth buffalo,
Ictiobus bubaZus
(Rafinesque), with reference to Elephant
Butte Lake. New Mexico State Univ., Las Cruces. Ag. Exp. Sta. Res. Rept.
261.?
111 p.
111
FISH TEMPERATURE DATA
Species:
?
Sockeye salmon,
Oncorhynchus nerka
I. Lethal threshold:
referencei
acclimation
temperature
larvae
?
juvenile
?
adult
Upper
5
22
1.
10
23
1
15
24
1
20
25
1
Lower
5
0
1
10
3
1
15
4
1
20
5
1
23
7
1
II. Growth:
larvae
jizerjile?
adult
Optimum and
15(5)
15(2)*
2,5
[range]
(10-15)
4
(11-17)
*Max. with excess food
III.
Reproduction:
optimum
range?
month(s)
Migration
7-16
4
Spawning
7-13?
Fall
6
Incubation
and hatch
acclimation
IV. Preferred:
?
temperature larvae
?
juvenile adult
Summer?
15
I
References on following page.
3
112
Sockeye salmon
References
1.
Brett, J. R. 1952. Temperature tolerance in young Pacific salmon, genus,
Oncorhynchus.
J. Fish. Res. Bd. Canada. 9:265-323.
2.
Griffiths, J. S., and D. F. Alderdice. 1972. Effects of acclimation and
acute temperature experience on the swimming speed of juvenile coho salmon.
J. Fish. Res. Bd. Canada. 29:251-264.
3.
Ferguson, R. G. 1958. The preferred temperature of fish and their mid-
summer distribution in temperate lakes and streams. J. Fish. Res. Bd.
Canada. 15:607-624.
4.
Burrows, R. E. 1963. Water temperature requirements for maximum productivity
of salmon. Proc. 12th Pacific N.W. Symposium on Water Poll. Res.., Nov. 7, 1963,
Corvallis, Oregon. pp. 29-32.
5. Shelbourn, J. E., J. R. Brett, and S. Shirahata. 1973. Effect of temperature
and feeding regime on the specific growth rate of sockeye salmon fry
(Oncorhynchus nerka)
with a consideration of size effect. J. Fish. Res. Bd.
Canada.
?
30:1191-1194.
6.
U. S. Environmental Protection Agency. 1971. Columbia River thermal effects
study, Vol. 1. Biological Effects Studies. 95.p.
7.
Donaldson, L. R., and F. J. Foster. 1941. Experimental study of the effects
of various water temperatures on growth, food utilization and mortality of
fingerling sockeye salmon. Trans. Amer. Fish. Soc. 70:339-346.
113
FISH TEMPERATURE DATA
Species:
Striped bass,
Morone saxatilis
I. Lethal threshold:
Upper
Lower
acclimation
temperature
larvae
?
juvenile?
adult
reference'
not given
35*?
28**
2
*Laboratory
**Field observation
II. Growth:
?
larvae
?
juvenile?
adult
Optimum and
[range]
III. Reproduction:
optimum
range
month(s)
Migration
6-8
2
Spawning
16-19(2)
12-22(1)
Apr-June(1)
1,2
16-24
Incubation
and hatch
acclimation
IV. Preferred:
?
temperature?
larvae
juvenile
?
adult
5 Dec
12
14 Nov
22
3
21
Oct
26
3
28
July
28
3
References on following page.
114
Striped bass
References
1.
Shannon, E. H. 1970. Effect of temperature changes upon developin
striped bass eggs and fry. Proc. 23rd Conf. S.E. Assoc. Game and
Fish Comm., October 19-22, 1969. pp. 265-274.
2.
Talbot, G. B. 1966. Estuarine environmental requirements and limiting
factors for striped bass. In: A Symposium on Estuarine Fisheries.
Amer. Fish. Soc., Special Publ. No. 3. pp. 37-49.
3.
Meldrim, J. W., J. J. Gift, and B. R. Petrosky. 1974. Supplementary
data on temperature preference and avoidance responses and shock
experiments with estuarine fishes and macroinvertebrates. Ichthyological
Associates, Inc., Middletown, Delaware. 56 p. mimeo.
115
FISH TEMPERATURE DATA
Species:
?
Threadfin shad,
Dorosoma petenense
acclimation
I.
Lethal threshold: temperature
?
larvae?
juvenile
?
adult
Upper
Lower?
9*
*lowest permitting
some survival
II. Growth:
?
larvae?
juvenile
?
adult
Optimum and
[range]
reference'
III. Reproduction:
Migration
?
-
optimum?
range
month(s)
Spawning
14(3)-23(4)
Apr-Aug(4)
3,4
Incubation
and hatch
23(4)-34(5)
4,5
acclimation
IV.
Preferred:
?
temperature larvae
?
juvenile?
adult
>19
References on following page.
116
Threadfin shad
References
1.
Strawn, K. 1963. Resistance of threadfin shad to low temperatures,
Proc. 17th Ann. Conf. S.E. Assoc. Game and Fish Comm., 1962. pp. 290-293.
2.
Adair, W. D., and D. J. DeMont. 1970. Effects of thermal pollution
upon Lake Norman fishes. N. Carolina Wildlife Res. Comm., Div. Inland
Fisheries, Raleigh, North Carolina. Summary Report, Fed. Aid Fish
Restoration Project F-19-2. 14 p.
3.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol.
1. Life History Data on Freshwater Fishes of the United States and
Canada, Exclusive of the Perciformes. Iowa State Univ. Press, Ames,
Iowa. 752 p.
4.
Shelton, W.' L. 1964. The threadfin shad,
Dorosoma petenense
(Gunther):
Oogenesis, seasonal ovarian changes and observations in life history.
M.S. Thesis, Oklahoma State Univ., Norman. 49 p.
5.
Hubbs, C., and C. Bryan. 1974. Maximum incubation temperature of the
threadfin shad,
Dorosoma petenense.
Trans. Amer. Fish. Soc. 103:369-371.
117
FISH TEMPERATURE DATA
Species:
Walleye,
Stizostedion vitreum
I. Lethal threshold:
acclimation
temperature?
larvae
juvenile
?
adult
reference
Upper
12
29
1
16
31
1
22
31
1
Lower
26
31
1
II. Growth:
Optimum and
[range]
larvae
juvenile
adult
22(1)
20(6)
(16-9s)
1
III. Reproduction:
optimum
range
month(s)
Migration
3-7
4
Spawning
6-9(1)*
4(7)-17(5)
Apr-May(4)
1,5,7,4
9-15
Incubation
and hatch
*for fertilization
acclimation
IV.?
Preferred:
temperature larvae?
juvenile adult
23*
2
22-25(1)
25(3)*
1,3
*field
References on following page.
118
Walleye
References
1.
Smith, L. L., Jr., and W. M. Koenst. 1975. Temperature effects on eggs
and fry of percoid fishes. U. S. Environmental Protection Agency, Duluth
Minnesota. Report EPA-660/3-75-017. 91 p.
2. Ferguson, R. G. 1958. The preferred temperature of fish and their mid-
summer distribution in temperate lakes and streams. J. Fish. Res. Bd.
Canada. 15:607-624.
3.
Dendy, J. S. 1948. Predicting depth distribution of fish in three TVA
storage reservoirs. Trans. Amer. Fish. Soc. 75(1945):65-71.
4. Eddy, Samuel, and T. -Surber. 1943. Northern Fishes with Special Reference
to the Upper Mississippi
.
Valley. Univ. Minn. Press, Minneapolis. 276 p.
5.
Niemuth, W., W. Churchill, and T. Wirth. 1959. The walleye, its life
history, ecology, and management. Wisc. Cons. Dept., Madison. Pub. 227.
14 p.
6.
Kelso, John R. M. 1972. Conversion, maintenance, and assimilation for
walleye,
Stizostedion vitreum vitreum,
as affected by size, diet, and.
temperature. J. Fish. Res. Bd. Canada. 29:1181-1192.
7.
Grimstead, Bobby G. 1971. Reproduction and some aspects of the early
life history of walleye,
Stizostedion vitreum
(Mitchell), in Canton
Reservoir, Oklahoma. In: Reservoir Fisheries and Limnology. Amer.
Fish. Soc., Washington, D. C. Spec. Pub. No. 8. G. Hall, ed. pp. 41-51.
119
FISH TEMPERATURE DATA
Species:
White bass,
Morons chrysops
acclimation
I. Lethal threshold: temperature larvae?
juvenile?
adult
Upper
Lower
17?
14*
*% mortality not given
II.
Growth:
?
larvae?
juvenile
?
adult
Optimum and
?
24-30
[range]
III.
Reproduction:
?
optimum?
range
?
month(s)
Migration
Spawning
Incubation
and hatch
14-20 (north)
12-?
,
(Tenn) .Mar-May Tenn
16(2)-26(6)
reference'
3
7
4
1
2,6
acclimation
IV. Preferred:?
temperature larvae
?
juvenile adult
Summer
28-30*
*Field
References on following page.
120
White bass
References
1.
Webb, J. F., and D. D. Moss. 1967. Spawning behavior and age and growth
of white bass in Center Hill reservoir, Tennessee. M.S. Thesis, Tenn.
Tech. Univ.
2.
Yellayi, R. R. 1972. Ecological life history and population dynamics
of white bass,
Morone chrysops
(Rafinesque) in Beaver Reservoir. Part
2. A contribution to the dynamics of white bass,
Morone chrysops
(Rafinesque)
population in Beaver Reservoir, Arkansas. Report to Arkansas Game and Fish
Commission. Univ. of Arkansas., Fayetteville.
3. Duncan, T. 0., and M. R. Myers. Artificial rearing of white bass,
Roccus
chrysops,
Rafinesque. Unpublished data. South Central Reservoir Inves-
tigations, Bureau Sport Fisheries •and Wildlife, Fayetteville, Arkansas.
4.
Ruelle, R. 1971. Factors influencing growth of white bass in Lewis and
Clark Lake. In: Reservoir Fisheries and•Limnology. Amer. Fish. Soc.
Washington, D. C. Spec. Pub. No. 8. G. Hall, ed., pp. 411-423.
5. Gammon, J. R. 1973. The effect of thermal input on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Research Center, Lafayette, Indiana. Tech. Rept. 32. 106 p.
6.
McCormick, J. H. 1976. Temperature effects on white bass
(Morone chrysops)
embryo development, and survival of one-day-old larvae. U. S: Environmental
Protection Agency, Duluth, Minnesota. In preparation.
7.
McCormick, J. H. 1976. Temperature effects on the growth of juvenile white
bass. U. S. Environmental Protection Agency, Duluth, Minnesota. In
preparation.
121
FISH TEMPERATURE DATA
Species:
White crappie,
Pomoxis annularis
acclimation
I. Lethal threshold: temperature
Upper?
29
larvae
juvenile
?
adult
33
reference'
4
Lower
II. Growth:
Optimum and
[range]
larvae
juvenile
25
adult
4
III. Reproduction:
Migration
optimum
range
month(s)
Spawning
16-20(5)
14-23(5)
_Mar-July(3) 3.5
19
?
_
14-23
5
Incubation
and hatch
Hatch in 24-27-1/2 hrs. at 21-23
acclimation
2
IV.?
Preferred:
temperature
larvae
?
juvenile
adult
27?
July(6)
28(6) 28-29(1) 1,6
3?
Jan
8
6
5?
Mar
10
6
24
?
June
26
6
References on following page.
122
White crappie
References
1. Gammon, J. R. 1973. The effect of thermal input on the populations of
fish and macroinvertebrates in the Wabash River. Purdue Univ. Water
Resources Research Center, Lafayette, Indiana. Tech. Rept. 32. 106 p.
2.
Breder, C. M., and D. E. Rosen. 1966. Modes of Reproduction in Fishes.
Nat. History Press, Garden City, New York. 941 p.
3. Goodson, Lee F. 1966. Crappie. In: Inland Fisheries Management, A.
Calhoun, ed. Calif. Dept. Fish and Game. pp. 312-332.
4. Kleiner, C. F., and K. E. F. Hokanson. Effects of constant temperature
on growth and mortality rates of juvenile white crappie,
Pomoxix annularis
Rafinesque. Unpublished data, U. S. Environmental Protection Agency,
?
.
Duluth, Minnesota.
5.
Siefert, R. E. 1968. Reproductive behavior, incubation, and mortality
of eggs and post larval food selection in the white crappie. Trans.
Amer. Fish. Soc. 97:252-259.
6.
Peterson, S. E., R. M. Schutsky, and S. E. Allison. 1974. Temperature
preference, avoidance, and shock experiments with freshwater fishes and
crayfishes. Ichthyological Associates, Inc., Drumore, Pennsylvania.
Bulletin 10.
123
FISH TEMPERATURE DATA
Species:
White perch,
Morone americana
acclimation
I.
Lethal threshold: temperature larvae
?
juvenile?
adult
Upper
Lower
II.
Growth:?
larvae?
juvenile
?
adult
Optimum and
[range]
reference'
III. Reproduction:
optimum
range
month(s)
Migration
Spawning
11(3)-20(1)
May-June(3)
1,3
Incubation
and hatch
acclimation
IV.?
Preferred:
temperature
larvae
?
juvenile adult
6
10
2
15
20
2
20
25
2
26-30
31-32
2
' References on following page.
124
White perch
References
1.
Holsapple, J. G., and L. E. Foster. 1975. Reproduction of white perch in
the lower Hudson River. New York Fish and Game J. 22:122-127.
2. Meldrim,
J. W., J. J.
Gift, and B. R. Petrosky. 1974. Supplementary data
on temperature preference and avoidance responses and shock experiments
with estuarine fishes and macroinvertebrates. Ichthyological Associates,
Inc., Middletown, Delaware. 56 p. mimeo.
3.
Sheri, A. N., and G. Power. 1968. Reproduction of white perch,
Roccus
americana,
in the bay of Quinte, Lake Ontario. J. Fish. Res.
•
Bd. Canada.
25:2225-2231.
FISH TEMPERATURE DATA
Species: ?
White sucker,
Catostomus commersoni
acclimation
I. Lethal threshold: temperature larvae
?
juvenile?
adult
5?
26''
Upper?
1
0 ?
28(11*?
28! 221
15. ?
31(1)?
29(2)
20(2), 21(1)?
30(1)?
29(2)
25
?
29
.25-26
?
31.
Lower
?
*7-day
TL50
for swimup
?
2h?
2-1
21 ?
6*
25?
6
*7-day TL50 for swimup
II. Growth:?
larvae?
juvenile
?
adult
Optimum and
?
27
[range]
?
(24-27)
III.
Reproduction:?
optimum
?
range
?
month(s)
Migration
Spawning
q,10(5)
?ral.?
-18(5,6)?
Mar-June(2)
incubation
and hatch?
15
?
9-20
acclimation
IV. Preferred:
?
temperature larvae
?
juvenile adult
19-21
reference
1,2
1,2
2
3
1
2,5,6
References on:
following page.
126
White sucker
References
1.
McCormick, J. H., B. R. Jones, and K. E. F. Hokanson. 1976. Temperature
effects on embryo development, early growth, and survival of the white
sucker,
Catostomus commersoni
(Lacepede). J. Fish. Res. Bd. Canada. In
Press.
2.
Carlander, K. D. 1969. Handbook of Freshwater Fishery Biology, Vol. 1.
Life History Data on Freshwater Fishes of the United States and Canada,
Exclusive of the Perciformes. Iowa State Univ. Press, Ames Iowa. 752 p..
3.
Brett, J. R. 1944. Some lethal temperature relations of Algonquin Park
• fishes. Pubi. Ont. Fish. Res. Lab. 63:1-49.
4.
Horak, D. L., and H. A. Tanner. 1964. The use of vertical gill nets in
studying fish depth distribution: Horsetooth Reservoir, Colorado. Trans.
Amer. Fish. Soc. 93:137-145.
5.
Webster, D. A. 1941. The life history of some Connecticut fishes. In:
A Fishery Survey of Important Connecticut Lakes. Conn. Geol. and Nat.
Hist. Survey Bull. No. 63, Hartford. pp. 122-227.
6.
Raney, E. C. 1943. Unusual spawning habitat for the common white sucker
Catostomus c. commersonii.
Copeia. 4:256.
127
FISH TEMPERATURE DATA
Species:
Yellow perch,
Perca flavescens
acclimation
I. Lethal threshold:
Upper
Lower
temperature
larvae
juvenile
adult
5
21
1
10(1),
10(4)
10(4)*
25(1)
1,4
15(1),
20(4)
19(4)*
28(1)
1,4
25
32
10
25
*swimup
9
10
II. Growth:
larvae
juvenile
adult
Optimum and
28
11.
[range]
(26
- 3
0
)(
1
1)
[13(6)-20(7)1
6,7,11
HI. Reproduction:
?
optimum
range.
month(s)
Migration
Spawning
?
12(3)
2(5)-15(3)
Mar-June(3)
3,5
Incubation
and hatch
?
10 up 1°/day
7-20
4
to 20
acclimation
IV. Preferred:?
temperature
larvae
?
juvenile
adult
Winter
21(2)
Summer
24
2
24
20-23
18-20
9
25
22
8
7
19
8
2
I
References on following page.
20
8
reference'
128
Yellow perch
References
1.
Hart, J. S. 1947. Lethal temperature relations of certain fish of the
Toronto region. Trans. Roy. Soc. Can., Sec. 5. 41:57-71.
2.
Ferguson, R. G. 1958. The preferred temperature of fish and their mid-
summer distribution in temperate lakes and streams. J. Fish. Res. Bd.
Canada. 15:607-624.
3. Jones, B. R., K. E. F. Hokanson, and J. H. McCormick. 1976. Winter
temperature requirements for maturation and spawning of yellow perch,
Perca flavescens
(Mitchell). U. S. Environmental Protection Agency,
Duluth, Minnesota. In preparation..
4.
Hokanson,
K.
E. F., and C. F. Kleiner. 1973. The effects of constant and
rising temperatures on survival and developmental rates of embryonic and
larval yellow perch,
Perca flavescens
(Mitchell). In: Early Life History
of Fish. Proceedings of an International Symposium, May 17-23, 1973,
Dunstaffnage Marine Research Lab., Oban, Scotland. pp. 437-448.
5. Muncy, R. J. 1962. Life history of the yellow perch,
Perca f7avescens,
in estuarine waters of Severn River, a tributary of Chesapeake Bay,
Maryland. Chesapeake Sci. 3:143-159.
6.
Coble, D. W. 1966. Dependence of total annual growth of yellow perch on
temperature. J. Fish. Res. Bd. Canada. 23:15-20.
7.
Weatherley, A. H. 1963. Thermal stress and interrenal tissue in the perch,
Perca fluviatilus
(Linnaeus). Proc. Zool. Soc., London. 141:527-555.
8.
Meldrim, J. W., J. J. Gift, and B. R. Petrosky. 1974. Supplementary data
on temperature preference and avoidance responses and shock experiments with
estuarine fishes and macroinvertebrates. Ichthyological Associates, Inc.,
Middletown, Delaware. 56 p. mimeo.
9.
McCauley, R. W., and L. A. A. Read. 1973. Temperature selections by juvenile
and adult yellow perch
(Perca flavescens)
acclimated to 24 C. J. Fish. Res.
Bd. Canada. 30:1253-1255.
10.
Hart, J. S. 1952. Geographic variations of some physiological and morphologi-
cal characters in certain freshwater fish. Univ. Toronto, Toronto, Ontario.
Biology Series No. 60. 78 p.
11.
McCormick, J. H. 1976. Temperature effects on growth of young yellow perch,
Perca flavescens
(Mitchell). U. S. Environmental Protection Agency, Duluth,
Minnesota. Report EPA-600/3-76-057.
129
(Please read
TECHNICAL
Instructions on
REPORT
the reverse before
DATA
completing)
1. REPORT NO.
EPA-600/3-77-061
2.
3. RECIPIENT'S ACCESSION•NO.
4. TITLE AND SUBTITLE
TEMPERATURE CRITERIA FOR FRESHWATER FISH:
PROTOCOL AND PROCEDURES
5. REPORT DATE
May 1977 issuing date
6. PERFORMING ORGANIZATION CODE
7. AUTHOR(S)
William A. Brungs and Bernard R. Jones
8. PERFORMING ORGANIZATION REPORT NO.
9. PERFORMING ORGANIZATION NAME AND
Environmental Research Laboratory-Duluth,
Office of Research and Development
ADDRESS
MN
Agency
10. PROGRAM ELEMENT NO.
1BA608
ii.
CONTRACT/GRANT NO.
In-house
U.S. Environmental Protection
Duluth, Minnesota?
55804
12. SPONSORING AGENCY NAME.AND ADDRESS
Same as above?
-
.
13. TYPE OF REPORT AND PERIOD COVERED
Tn-hOltaR
14.
SPONSORING AGENCY CODE
EPA/600/03
,
15. SUPPLEMENTARY NOTES
?
.
16. ABSTRACT?
•
The evolution of freshwater temperature criteria is discussed as it
relates to standards development by regulatory agencies. ?
The present,
generally accepted philosophical approach to criteria development is explained
in detail and its use to protect various life.stages of fish is demonstrated
by selected examples.
?
Numerical criteria for survival, spawning, embryo
development, growth, and gamete maturation of fish species were calculated
and tabulated.
?
-
17.?
KEY WORDS AND DOCUMENT ANALYSIS
a.
?
DESCRIPTORS
b. IDENTIFIERS/OPEN ENDED TERMS
C.?
COSATI
Field/Group
Temperature
Fresh Water Fishes
Growth
Reproduction (biology)
Mortality
Water Pollution
Water Quality Criteria
Water Quality Standards
Temperature Requirements
Thermal Pollution
06 S
06 F
06 C
18. DISTRIBUTION STATEMENT
RELEASE TO PUBLIC
19. SECURITY CLASS
(This
Report)
UNCLASSIFIED
21. NO. OF PAGES
136
20. SECURITY CLASS
(This
page)
UNCLASSIFIED
22. PRICE
EPA Form 2220-1 (9-73)
...ten c tinicentucur
001l17141c
nccirc?
inn !Jr, S-I I
U.S. ENVIRONMENTAL PROTECTION AGENCY
Office of Research and Development
Technical Information Staff
Cincinnati, Ohio 45268
OFFICIAL BUSINESS
PENALTY FOR PRIVATE USE, $300
AN EQUAL
OPPORTUNITY
EMPLOYER
H. L. Giesecke, (PS) 0001
Librarian
Illinois EPA
2200 Churchill Rd.
Springfield, IL 62706
POSTAGE AND FEES PAID
U.S. ENVIRONMENTAL PROTECTION AGENCY
EPA-335
Special Fourth-Class Rate
Book
If your address is incorrect, please change on the above label;
tear off; and return to the above address.
If you do not desire to continue receiving this technical report
series, CHECK HERE O; tear off label, and return it to the
above address.
Back to top